 ACME SMART PUBLICATION
ACME SMART PUBLICATION
- Books Name
- A TEXT OF BIOLOGY - CLASS XII
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 12
- Subject
- Biology
SPECIATION AND ISOLATION
Speciation is the formation of one or more new species from an existing species.
The crucial episode in the origin of species occurs when the gene pool of a population is severed from other populations of the parent species and gene flow no longer occurs.
Speciation can take place in two modes based on the geographical relationship of a new species to its ancestral species.
1. When a population, formerly continuous in range, splits into two or more geographically isolated populations and form new species, the mode of speciation is called allopatric speciation.
This can happen by subdivision of the original population, when a geographical barrier, such as a creeping glacier, a land bridge (e.g., Isthmus of Panama) or ocean or mountain, cuts across a species range.
Alternatively, a small number of individuals may colonise a new habitat which is geographically separated from the original range.
Darwin's finches that formed separate species in the Galapagos islands and the Australian marsupials that radiated to form new species are its examples.
2. In the second speciation mode, a subpopulation becomes reproductively isolated in the midst of its parent population; this is sympatric speciation.
So, sympatric speciation is the formation of species within a single population without geographical isolation.
The usually quoted example of sympatric speciation comes from polyploidy, which is the multiplication of the normal chromosome number.
This can happen when chromosomes fail to segregate at meiosis or replicate without undergoing mitosis.
Concept Builder
(I) Multiplicative speciation (Cladogenesis) : It is the formation of two or more species from a single species, example, allopatric speciation, sympatric speciation. This can be gradual or abrupt.
(II) Fusion species : It is an allogenous transformation. Isolation mechanism may break down due to a mutation. The two species will interbreed and merge to form a single species.
(III) Phyletic speciation (Anagenesis) : It is autogenous transformation of a species with passage of time due to piling up of variations.
3. Species concept:
Species is the basic unit of classification.
The term was coined by John Ray (1693).
Most taxonomists define species as morphologically distinct and reproductively isolated natural population or group of populations where individuals resemble one another more closely than with members of other species, have a similar anatomy, karyotype and biochemicals, interbreed freely and form a genetically closed system. There are three basic concepts about the species.
4. Morphospecies concept:
It is the earliest concept of species.
Davis and Heywood (1963) have defined it as "assemblage of individuals with morphological features in common and separable from other such assemblages by correlated morphological discontinuities in a number of features."
However, the number of morphological characters chosen for separating species varies from taxonomist to taxonomist.
"Lumpers" will combine all the populations with broadly similar traits into a single species while "Splitters" will separate various populations with even minor morphological differences into distinct species.
5. Biological species concept:
Though first proposed by Buffon (1753), biological species concept was formulated by Mayr(1942).
According to it, a biospecies (biological species, Biological Species Concept) is sexually interbreeding or potentially interbreeding group of individuals which is reproductively isolated from other species and is therefore, separated from others by absence of genetic exchange.
Normally species are distinct from one another by both morphological traits and reproductive isolation.
However, sibling species are those distinct species which are almost identical morphologically but are distinct from each other due to absence of interbreeding, e.g., Drosophila pseudoobscura and D. persimilis.
Biological species concept is, therefore, mainly based on absence of cross fertilisation between members of two species.
Cross fertilisation tests carried out by taxonomists between individuals of morphological and geographically separated populations have, resulted in revision of species and grouping of many of them into single species, e.g., several species of North American sparrows as subspecies and races of a single song sparrow, Passarella melodia.
The only problem of using reproductive isolation is the absence of sexual reproduction in several organisms -prokaryotes, some protists, some fungi, some plants (e.g., commercial Banana) and animals.
Further, cross fertilisation experiments cannot be performed on such a large number of species that occur in varied geographical areas.
Reproductive isolation cannot be used as a criterion in case of fossils.
The living organisms and fossils can be grouped only on the basis of their morphology and biochemistry.
Mayr (1987) has named morphologically grouped asexual species as paraspecies while Ghiselin (1987) has named them pseudospecies.
6. Evolutionary species concept:
All evolutionary taxonomist have been in search of a proper definition of species which is basic unit of classification.
One such definition has been given by Simpson.
According to Simpson (1961) "an evolutionary species is a lineage (an ancestor-descendent sequence of population) evolving separately from others and with its own unitary evolutionary role and tendencies."
The concept stresses on evolutionary isolation with sexual isolation being its one aspect.
It is more dependent on differences which can be morphological, genetical, behavioural and ecological, to know evolutionary distance.
However, evolution does not occur simultaneously in all the traits.
Neither its rate nor direction (in which it is occurring) are the same.
7. Reproductive isolation may be defined as the existence of intrinsic barrier to the interbreeding in natural populations. Each of these intrinsic barriers is called a reproductive isolating mechanism. According to Mayr (1942), reproductive isolating mechanisms are the biological properties of individuals which prevent the interbreeding of naturally sympatric populations.
8. Reproductive isolation in the form of hybrid sterility is known since long. In the laboratory or in zoos, hybrids can be produced between species that do not interbreed in nature. Horses and donkeys are two different species; a hybrid, mule, is produced from the mating of a male donkey and a mare (female horse).
9. Similarly, mating between stallion (male horse) and female donkey results in a hybrid called hinny. Both mule and hinny are sterile.
10. There are examples of species, which can produce fertile hybrids in captivity. You might have heard about the famous 'tigons', a hybrid of African lioness (Panthera leo) and Asian tigers (Panthera tigris), which is fertile. No barrier to hybridisation between these species has evolved during their long isolation from each other Natural selection has not favoured a reduction in hybridisation for the simple reason that no hybridisation has been possible. Other examples of species that breed in captivity and produce fertile hybrids are mallard (a duck) and the pintail duck, the polar bear and the Alaskan brown bear and the platy and swordtail fishes. But these species do not interbreed at all in natural condition.
Concept Builder
BARRIERS TO HYBRIDISATION
Prezygotic Mechanisms: (Prevent mating or formation of zygote)
1. Ecological isolation : Two species live in different habitats and do not meet. (One may be living in fresh water and the other in the sea).
2. Temporal isolation : Breeding seasons or flowering time may be different in the two species.
3. Behavioural isolation: The males of one animal species are unable to recognise the females of another species as potential mates.
4. Mechanical isolation: The structural differences in genitalia of individuals belonging to different animal species interfere with mating.
5. Gametic isolation : The sperms and ova of different species of animals are unable to fuse. In plants, the pollen coming from a different species may be rejected by the stigma.
Postzygotic Mechanisms : A hybrid zygote is formed but it may not develop into a viable fertile adult.
1. Hybrid Inviability : Hybrid zygotes fail to develop. In plants, embryos arising from interspecific crosses abort.
2. Hybrid sterility : Hybrid adults do not produce functional gametes. (Mules and hinny are common examples in mammals. Several hybrid ornamental plants are sterile.)
3. Hybrid breakdown : The offspring of hybrids are inviable or infertile.
MODERN SYNTHETIC THEORY OF EVOLUTION
1. Evolution on the grand scale of geological time is called macroevolution.
2. Evolution at genetic level is called microevolution.
3. Studies of how individual traits evolve within natural populations provide powerful evidence that natural selection can be a powerful agent of microevolutionary change within species. The progressive change in allele frequencies within the population is micro-evolution.
4. Units of evolution is population.
5. Unit of natural selection is individual.
The modern synthetic theory of evolution is the result of the work of a number of scientists as T. Dobzhansky, R.A. Fisher, J.B.S. Haldane, Sewall Wright, Stebbins.
The synthetic theory includes the following factors.
(i) Gene mutations
(ii) Changes in chromosome structure and number
(iii) Genetic recombination
(iv) Natural selection
(v) Reproductive isolation
(vi) Migration lied
(vii) Hybridisation
NEUTRAL THEORY OF EVOLUTION
According to Kimura most of the mutations are neutral, and are not eliminated from the population.
This is against natural selection.
Kimura proposed that speciation is not due to selection of advantageous genotypes no but elimination of deleterious alleles and random selection of neutral alleles.
It emphasized that most mutations are of neutral value and genetic drift is responsible for divergence.
It means that all mutations are alike in adaptive value.
It is only chance or random drift which delineates a novel collection of mutants into a group divergent from the parental population.
Place of Humans in the Animal Kingdom
Today human evolution is being studied by:
(i) Homology in the chromosomes of man and great apes. The banding pattern of human chromosome number 3 and 6 are compared with those of particular autosomes in the chimpanzee. It shows a common origin for man and chimpanzee.
(ii) Today, besides the autosomal chromosomes, V-chromosomes and mitochondrial DNA are being studied, as they are uniparental in origin and do not take part in recombination.
(iii) Evidence from blood proteins -It has been proved by the blood protein tests that man is most closely related to great apes (chimpanzee and gorilla).
(iv) Evidence from blood groups -The blood groups A and B are found in apes and not in monkeys.
(v) Evidence from haemoglobin -There is 99 percent homology in haemoglobin of man and gorilla.
Human beings are vertebrates and belong to the class mammalia.
Mammals evolved from primitive reptiles in early Triassic period, about 210 million years ago.
But for nearly 150 million years, mammals existed as relatively inconspicuous group of small rat-like creatures, completely dominated by the large number of gigantic reptiles of the Mesozoic age.
It is only after the great extinction of dinosaurs and other large reptiles, that mammals diversified and began to occupy the earth's many different habitats.
Within the class mammalia, human beings belong to the order primates, a group that originated about 65 million years ago and includes not only monkeys and apes but also the lorises, lemurs and tarsiers.
The Anthropoid apes or the ancestors of monkeys, apes and humans evolved about 36 million years ago and the hominids or the ancestors of apes and humans evolved about 24 million years ago.
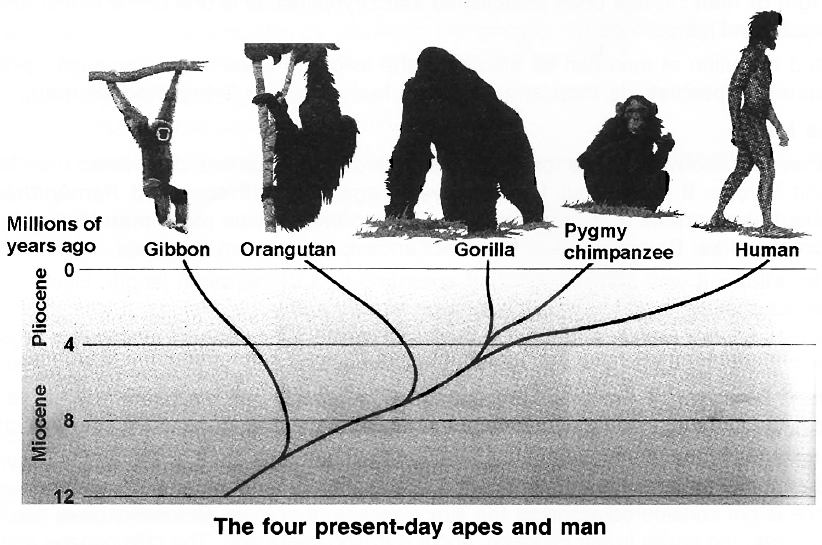
Today, the apes are represented by two families, namely, Pongidae which include chimpanzees, gorillas and orangutans and Hylobatidae which includes gibbons.
The chimpanzee and gorilla are restricted to Africa, whereas the orangutans and gibbons are found only in Asia.
Humans belong to the family Hominidae in which Homo sapiens is the only living species.
Early Human Ancestors
Tracing the evolution of human beings, both by fossil hunting and molecular methods, is one of the most exciting and active areas of research in biology.
The fossil evidence clearly indicated that such genera as Ramapithecus and Sivapithecus were the forerunners of Hominids.
A genus called Australopithecus appeared in Africa about five million years ago and ultimately gave rise to Homo about two million years ago.
But even Australopithecus had a brain measuring only about 350-450 cm3.
The most important change that must have occurred during the three million years or so, between the appearance of Australopithecus and that of our genus, must therefore have been a phenomenal increase in brain size, all the way up to 1400 to 1450 cm3, that is characteristic of our species.
A combination of molecular data and a modern interpretation of the fossil record suggests that the gibbons probably diverged from the main line of Hominoid evolution about ten million years ago, that the orangutan did so about eight million years ago and that the ancestors of gorilla and chimpanzee, about four million years ago.
The gorilla and chimpanzee have become separated from each other only 2.3 million years ago.
Place and Sequence of Human Evolution
There is evidence that almost all of Hominid evolution occurred in Africa, and Asia and that the evolution of the human species took place in Africa.
Several species belonging to the genus Homo can be recognised from the fossil record. For example, Homo habilis lived in Africa, about two million years ago and was characterised by having a larger brain than Australopithecus, used tools and was bipedal.
Another species, Homo erectus appeared about 1.7 million years ago, used fire and is believed to have migrated to Asia and Europe.
Fossils of the socalled 'Java man' and 'Peking man', belong to Homo erectus. Homo erectus was replaced by Homo sapiens.
A primitive form of Homo sapiens, called Neanderthal man (Homo sapiens neanderthalensis), was common in Europe and Asia.
The Neanderthal men resembled us, though they were relatively short and stocky and more powerfully built.
The Neanderthals made tools and used animal hides as clothing.
They built hut-like structures for dwelling and buried their dead.
There is evidence that an abrupt transition occurred all over Europe whereby the Neanderthal man was wiped out and gave way to the more efficient cousin, the cro-Magnon, about 34,000 years ago.
The Cro-Magnon people left behind very elaborate cave paintings showing the attainment of a form of culture not unlike our own.
After the last glacial period (about 10,000 years ago), modern Homo sapiens sapiens began to spread all over the globe, cultivated plants, domesticated animals and reached enormous population sizes.
Homo sapiens appeared in Africa about 500,000 years and probably replaced Homo erectus there.
But in Asia, Homo erectus appears to have survived for another 250,000 years when it was finally replaced by Homo sapiens migrating from Africa.
