 ACME SMART PUBLICATION
ACME SMART PUBLICATION
- Books Name
- A TEXT OF BIOLOGY - CLASS XII
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 12
- Subject
- Biology
MUTATION
Mutation is sudden, inheritable, discontinuous variation due to change in chromosomes and genes.
Hugo de Vries (1901) one of the discoverers of Mendel's laws, observed two distinct varieties of Oenothera lamarckiana (Evening primrose).
These differed in length of stem, flower form and colour and shape of leaves.
These mutant varieties are now known to have been produced due to chromosomal aberrations.
Seth Wright (1791) is considered to be the first to record point or gene mutation.
He noticed a lamb with unusually short legs.
This short legged breed of sheep was known as ancon breed.
Darwin called this variation as sports.
Bateson (1894) termed them as discontinuous or saltatory variations.
The credit for starting scientific study of mutations goes to Thomas Hunt Morgan (1910).
He is known for his work on fruit fly, Drosophila melanogaster.
He found white eyed mutant of Drosophila and since then about 500 mutations have been observed by geneticists from different parts of the world.
Types of Mutations:
Different classifications of mutations are known, each based on a definite criterion or character.
I. Spontaneous and induced mutations:
These are based upon the agency involved.
Spontaneous mutations. These are natural mutations. They have also been called background mutations. Such mutations occur at a frequency of 1 × 10–5 in nature.
Induced mutations. These have been observed in organisms due to specific factors such as radiations, ultra violet light or variety of chemicals. The agents which induce mutations on their application, are called mutagens or mutagenic agents.
II. On the basis of the type of cells in which mutations occur, there are other two types of mutations.
(a) Somatic mutations. These mutations occur in somatic cells, i.e., body cells or the cells other than germinal cells. The somatic mutations do not have any genetic or evolutionary importance. This is because only the derivatives or the daughter cells formed from the mutated cell will show mutation and not the whole organism.
(b) Germinal mutations. These mutations occur in the gametes or germ cells and are also known as gametic mutations. Such mutations are heritable, and, therefore, are of great evolutionary significance. If the mutations are dominant, these are expressed in the next generation and if recessive, their phenotypic expressions remain suppressed.
III. Forward and backward mutations:
The commonest type of mutation, is the change from normal or wild type to new genotype (recessive or dominant).
Such mutations are called forward mutations.
An organism which has undergone forward mutation, may again develop mutation which restores the original wild-type phenotype.
Such reversions are known as backward mutations or reverse mutations.
Mutations can occur at any stage during the life cycle of an organism.
Types of Mutations:
1. Gene mutation.
It is alteration in the sequences of nucleotide in nucleic acids or any change in the sequence of triplet bases.
If gene mutation arise due to change in single base pair of DNA, it is called point mutation.
Gene mutation occurs by following methods :
(a) Frame-shift mutation (Gibberish mutation)
(i) Deletion : Removal of one or more bases from nucleotide chain.
(ii) Insertion or addition: Addition of one or more bases in a nucleotide chain.
(b) Substitution. The replacement of one base by another.
It is of two types :
(i) Transition: When a purine base (A or G) is substituted by another purine base or pyrimidine base (T or C) is substituted by another pyrimidine base.
(ii) Transversion: Substitution of a purine base with a pyrimidine base or vice versa.
(c) Tautomerization: The purines and pyrimidines in DNA and RNA may exist in several alternate forms or tautomers. Tautomerization occurs through rearrangement of electrons and protons in the molecule. Tautomers show changed base pairing so as to cause change in sequence like AT to CG.
Non-sense Mutation : Such mutations arise when a normal codon, coding for an amino acid is changed into a chain terminating codon (UAG, UAA, UGA) resulting in the production of an incomplete polypeptide.
Nonsense mutations rarely go unnoticed because the incomplete or shorter protein formed, is generally inactive.
Mis-sense mutation: Change in base in a codon, producing a different amino acid at the specific site in a polypeptide. In mis-sense mutation, the change in one amino acid is frequently compatible with some biological activity, e.g., sickle cell anaemia.
2. Chromosomal mutation (Chromosomal Aberrations) :
The change in the chromosome morphology is called chromosomal aberration. Structural changes in the chromosomes take place during meiosis. There are four types of chromosomal rearrangements:
(a) Deficiency :
Loss of a terminal segment of chromosome or Deletion -It is due to loss of a intercalary part of chromosome, e.g., Cri-du-Chat syndrome (short arm of chromosome 5 loses a part).
(b) Duplication:
Occurs due to addition of a part of chromosome so that a gene or set of genes is represented twice, e.g., Barr eye in Drosophila.
(c) Translocation :
It involves shifting of a part of one chromosome to another non-homologous chromosome. So new recombinant chromosomes are formed, as this induces faulty pairing of chromosomes during meiosis. An important class of translocation having evolutionary significance is known as reciprocal translocation or segmental interchanges, which involves mutual exchange of chromosome segments between non-homologous chromosome, i.e., illegitimate crossing over. Chronic myelogenous leukemia (CML) occurs due to translocation of segment of long arm from chromosome 22 to chromosome 9. Chromosome 22 is called Philadelphia chromosome.
(d) Inversion:
Change in linear order of genes by rotation of a section of chromosome by 180°. Inversion occurs frequently in Drosophila as a result of X-ray irradiation. They may be of two types:
(i) Paracentric : Inversion without involving centromere (Inverted segment does not carry centromere).
(ii) Pericentric: Inversion involving centromere.
3. Genomatic mutation or numerical changes in chromosome number :
It is of two types:
(A) Aneuploidy:
In aneuploidy, any change in number of chromosomes in an organism would be different than the multiple of basic set of chromosomes. It results due to failure of segregation of chromatids during cell division cycle. There would be following two possibilities:
(a) Hypoploidy:
This arises due to loss of one or more chromosomes or pairs of chromosome. Thus, the following conditions are likely to be produced:
(i) Monosomy (2n -1) : It is the result of loss of one chromosome for a homologous pair. Its other variant is double monosomy, i.e., (2n -1 -1).
(ii) Nullisomy (2n -2) : It is the result of loss of a complete homologous pair of chromosome.
(b) Hyperploidy: This arises due to addition of one or more chromosomes or pairs of chromosome. The following conditions are thus likely to be produced :
(i) Trisomy (2n + 1) : In this type, a single chromosome is added to the chromosome set. The trisomics were obtained for the first time in Datura stramonium (Jimson weed) by A.F. Blakeslee and his co-workers (1924). In human beings, Mongolism or Down's syndrome is due to trisomy of chromosome 21 (2n + 1 or 46 + 1, i.e., chromosome 21 is present 3 times). Others are Patau syndrome (due to trisomy of 13th) and Edward's syndrome (due to trisomy of 18th chromosome).
(ii) Tetrasomy (2n + 2) : It is the result of addition of a complete homologous pair of chromosomes (i.e., 2-chromosomes).
(B) Euploidy:
In euploidy, any change in the number of chromosomes is the multiple of the number of chromosomes in a basic set or it occurs due to variation in one or more haploid sets of chromosomes. Accordingly, these may be haploid and polyploid.
(a) Haploidy : One set of chromosomes. Haploids are better for mutation experimental studies, because all mutations either dominant or recessive can express immediately in them, as there is only one allele of each gene present in each cell.
(b) Polyploidy: Failure of cytokines is after telophase stage of cell division results in an increase in a whole set of chromosomes in an organism and this phenomenon is known as polyploidy. Polyploids are of particular importance and are, therefore, discussed below. Polyploids fall into two major categories:
(i) Autopolyploids have the same basic set of chromosomes multiplied more than twice, e.g., AAA -autotriploid, AAAA -Autotetraploid. Autopolyploids with odd number of chromosomes are seedless but show Gigantism (large size) or gigas effect (more yield and more adaptability), but are odd numbered so can only be propagated vegetatively, e.g., Banana, Pineapple. Naturally occurring autotriploids are known in banana, grapes, sugar beet, tomato and watermelons. Similarly, autotetraploids occur amongst apples, berseem, corn etc.
Autopolyploids can also be produced artificially by treating the seeds, seedlings or axillary buds with an alkaloid called colchicine. It is extracted from corm of Colchicum autumnale. The treatment produces doubling of chromosomes.
(ii) Allopolyploids are hybrids whose chromosome sets are derived from two different genomes.
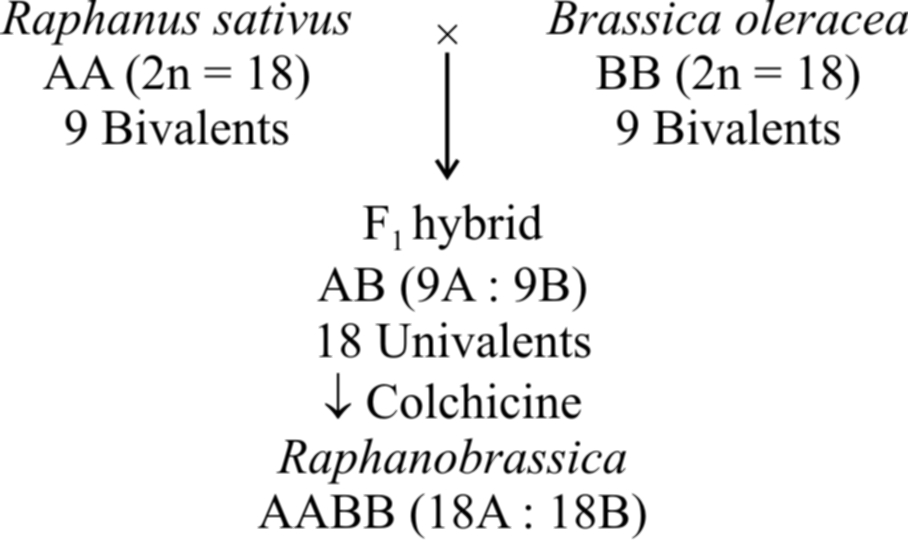
Let A and B be considered as different genomes. The two diploid organisms should have AA and BB chromosome sets. The autotetraploid can now be represented as AAAA while allotetraploid is represented as AABB.
Such polyploids are the result of doubling of chromosomes in F1 hybrids derived from two different or related species.
The commonest example of allopolyploidy is Raphanobrassica developed by Russian geneticist G.O. Karpechenko (1927). A cross was made between Brassica oleracea (2n = 18) and Raphanus sativus (2n = 18). F1 hybrid produced was sterile, because chromosome sets of both these plants were dissimilar and could not pair during meiosis. However, some fertile plants with 2n = 18A + 18B (18 bivalents) were found amongst these.
Another example is a man made cereal -Triticale, produced by (i) crossing Triticum durum (2n = 28) with Secale cereale (2n = 14) and then treating F1 hybrid with colchicine to obtain hexaploid Triticale and (ii) crossing Triticum aestivum (2n = 42) with Secale cereale (2n = 14) and then treating F 1 hybrid with colchicine to produce octaploid Triticale.
Mutagens
Mutations can be artificially produced by certain agents called mutagens or mutagenic agents. Following are two major types of mutagens :
(1) Physical mutagens
(2) Chemical mutagens
(1) Physical mutagens:
Radiations are the most important physical mutagens.
H.J. Muller who used X-rays, for the first time, to increase the rate of mutation in Drosophila, opened an entirely new field in inducing mutations.
So, Muller is considered as father of Actinobiology.
The main source of spontaneous mutations are the natural radiations coming from cosmic rays of the sun.
The spectrum of wavelengths that are shorter (i.e., of higher energy) than the visible light can be subdivided into following two groups.
(a) Ionizing radiations
(b) Non-ionizing radiations
They occur in small amounts in the environment and are known as background radiations.
The following are biological effects of radiations :
(a) Effects of ionizing radiations: The ionizing radiations include X-rays, -rays, -rays and -rays. Ionizing radiations cause breaks in the chromosome. These cells then show abnormal cell divisions. If these include gametes, they may be abnormal and even die prematurely. Different types of cancers may result due to radiations. The frequency of induced mutations is directly proportional to the doses of radiations.
(b) Effects of non-ionising radiations : The non-ionizing radiations have longer wavelengths but carry lower energy. This energy is insufficient to induce ionization. Therefore, non-ionizing radiations such as UV light do not penetrate beyond the human skin . Thymine (pyrimidine) dimer formation is a major mutagenic effect of UV rays that disturbs DNA double helix and thus, DNA replication.
(2) Chemical mutagens:
Large number of chemical mutagens are now known. These are more injurious than radiations. The first chemical mutagen used was mustard gas by C. Auerbach et. al. during world war II.
Chemical mutagens are placed into two groups:
(a) Those which are mutagenic to both replicating and non-replicating DNA such as nitrous acid,
(b) Those which are mutagenic only to replicating DNA, such as acridine dyes and base analogues.
Following are the effects of some of the chemical mutagens:
(i) Nitrous acid: It is mutagenic to both replicating and non-replicating DNA. It acts directly by oxidative deamination on A, G and C bases which contain amino groups. A is deaminated to hypoxanthine which is complementary to cytosine. G is converted to xanthine which pairs with C. Cytosine is converted to U which pairs with A
(ii) Acridines: Acridines and proflavins are very powerful mutagens. These can intercalate between DNA bases and interfere with DNA replication, producing insertion or deletion or both of single bases respectively. This induces frame shift-mutations or gibberish mutation, e.g., Thalassaemia.
(iii) Base analogues: These have structures similar to the normal bases and are incorporated into DNA only during DNA replication. Base analogues cause mispairing and eventually give rise to mutations. The base analogues may either be natural or artificial. Natural base analogues include 5-methyl cytosine, 5-hydroxymethyl cytosine, 6-methyl purine etc.
The most commonly used artificial base analogues are 5-bromouracil and 2-aminopurine. 5-bromouracil is a structural analogue of thymine. It gets incorporated into the newly replicated DNA in place of thymine (T). 2-aminopurine is an artificial base analogue of adenine. It acts as a substitute of adenine (A) and can also pair with cytosine (C).
CYTOPLASMIC INHERITANCE
Some self replicating genes (DNA) are present in the cytoplasm (mitochondrial DNA and chloroplast DNA) also.
These are called plasmagenes and all the plasmagenes together constitute plasmon (like genome).
The inheritance of characters by plasmagenes is called extranuclear or extrachromosomal inheritance.
Certain most important examples of extranuclear inheritance in eukaryotes are following :
Maternal inheritance:
The amount of nuclear hereditary material contributed by the two sexes is almost equal but the cytoplasm in egg is always much more than that of the sperm.
So, in extranuclear inheritance, contribution of female parent is more.
This is called maternal inheritance.
The evidence of maternal inheritance is the coiling of shell in snails.
Concept Builder
Shell Coiling in Snails
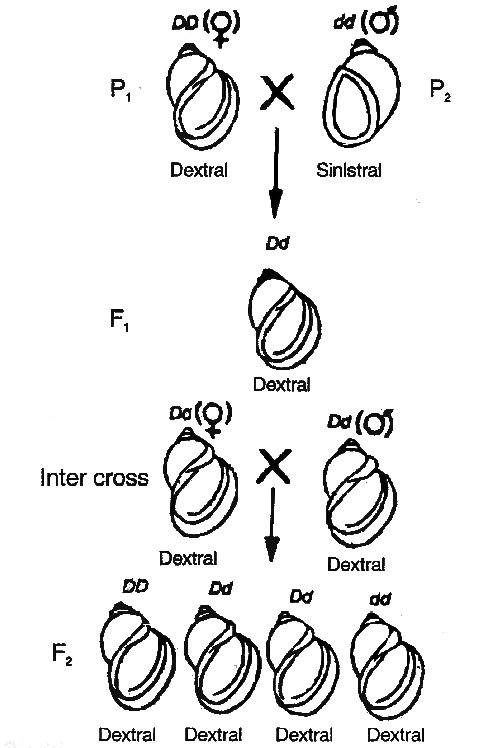
Shell coiling in Limnaea peregra, a fresh water snail, is of two types, dextral ( =clockwise) and sinistral (anticlockwise).
 dextral (DD) and
dextral (DD) and  sinistral (dd) snails
sinistral (dd) snailsDextral coiling is controlled by a dominant nuclear gene called D while sinistral coiling is due to a recessive gene d.
Since, zygote receives whole of its cytoplasm from the egg, the direction of shell coiling in the offspring is governed by cytoplasm of the mother.
When dextral female (DD) is crossed with sinistral male (dd), all the offsprings of F1 generation (Dd) have dextral coiling.
If sinistral female (dd) is crossed with dextral male (DD), the offspring have Dd genotype but coiling is sinistral.
When the latter are self-bred, three types of genotypes are produced1DD, 2Dd and 1dd.
However, shell coiling in all is dextral because the mother has Dd genotype.
Organelle inheritance:
The DNA is present in mitochondria and chloroplast which controls the inheritance of some characters.
A well known example of the characters controlled by chloroplasts is plastid inheritance in Mirabilis jalapa (4 O'clock plant), discovered by Correns.
Other examples of organellar inheritance are iojap inheritance in maize, inheritance of poky (imbalance in the mitochondrial physiology) in the fungus Neurospora crassa and Petite in yeast, a mitochondrial character.
Cytoplasmic male sterility in maize, is also a function of defective mitochondria.
