 ACME SMART PUBLICATION
ACME SMART PUBLICATION
Mendel's Laws of Inheritance
- Books Name
- A TEXT OF BIOLOGY - CLASS XII
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 12
- Subject
- Biology
INHERITANCE: HEREDITY AND VARIATIONS
Heredity :
It is the transmission of genetic characters from parents to the offsprings.
It deals with the phenomenon of "like begets like", e.g., human babies are like human beings in overall characteristics.
About 200 characters are found to be hereditary in man.
Variations are common in sexually reproducing organisms.
Asexually reproducing organisms are monoparental, hence exhibit no genetic variations.
Concept Builder
Variations are of following two types:
(1) Somatogenic Variations: These are acquired variations and are non-inheritable in nature. The ability of an organism to alter its phenotype in response to environment is called phenotypic plasticity.
(2) Blastogenic Variations: These are germinal variations and are hereditary in nature. They are again of two types :
(a) Continuous Variations: These are the fluctuating variations and can not give rise to new species. These are further of two types :
(i) Substantive: Variation in size, shape and colour of organism.
(ii) Meristic: Variation in number of parts, e.g., number of grains in an ear of wheat.
(b) Discontinuous Variations: Also known as mutations, sports or saltations. These variations are responsible for formation of new species and the organism thus formed, is called' a mutant.
Types of discontinuous variations :
(i) Substantive variations: These influence shape, colour, size etc., e.g., hairless cat, short legged ancon sheep.
(ii) Meristic variations: These affect number of parts, e.g., polydactyly in humans. Variations are of great significance in evolution as they make the organism better suited to modifying environmental conditions, produce new trait in organism and provide raw material for evolution.
Branches of Genetics
(i) Transmission genetics or Classical genetics, e.g., It is the study of Mendelian genetics and non-Mendelian genetics.
(ii) Forward genetics : It is the identification of mutated gene using the mutated phenotype.
(iii) Reverse genetics : It is the study of genes whose protein products are unknown.
(iv) Cytogenetics: It is the study of various aspects of chromosomes.
(v) Molecular/biochemical genetics : It is the study of structure and functions of genes.
(vi) Population or biometrical genetics : It is the study of the behaviour and effects of gene in population using mathematical models.
(vii) Behavioural genetics: It is the study of interaction of genes with the environment to produce a particular pattern of behaviour.
Pre-Mendelian Ideas About Inheritance or Theories of Blending Inheritance
The science of genetics arose with the rediscovery of Mendelism in 1900. Early philosophers, thinkers and workers have presented various theories to explain the phenomenon of inheritance.
These are called theories of blending inheritance. Some of these theories are:
(a) Moist Vapour Theory (Pythagoras : 500 B.C.) : Various body parts emit certain vapours, which get aggregated to form new individual.
(b) Reproductive Blood Theory (Aristotle: 384-322 B.C.) : According to him, menstrual fluid and semen are kinds of highly purified blood. Menstrual fluid provide inert substance for embryo formation and semen provide form and shape to embryo.
(c) Preformation Theory or Homunculus theory (J. Swammerdam) : According to this theory miniature form of individual is already present in sperm or egg called "homunculus". Fertilization is required to stimulate its growth.
(d) Theory of Pangenesis (Darwin 1868) : According to him, each part of body produces minute particle called gemmules or pangenes, which aggregate into gamete. On fusion they give rise to new individual.
(e) Theory of Epigenesis (K.F. Wolff) : According to this idea,· neither egg nor sperm had a structural homunculus but the gametes contained undifferentiated living substance capable of forming the organized body after fertilization. This suggested that many new organs and tissue which were originally absent, develop structurally de novo due to mysterious vital force.
Theory of pangenes was disproved by Weismann.
A.Weismann proposed his theory of germplasm, according to which the changes which affect the germplasm are heritable and the changes which affect somatoplasm are non heritable.
Objections to blending inheritance:
1. Unisexual traits
2. Skin colour in humans
3. Atavistic character
GENETIC TERMS AND SYMBOLS
MENDELIAN INHERITANCE
Mendel was born on July 22, 1822. He worke
d on Pisum sativum (Garden pea or Edible pea) for 7 years by taking 7 pairs of contrasting traits.
The results were read out in two meeting of Natural History Society of Brunn in 1865.
His paper "Experiments on plant Hybridisation" was published in 4th volume of "Proceedings of Natural Science Society of Brunn" in 1866.
Mendel was first to apply statistical analysis and mathematical logic.
He selected 14 true breeding pea plant varieties. He died due to kidney disorder in 1884.
Mendel selected following characters in pea plant for carrying out hybridisation experiments:

Mendel failed to produce same results in Hawkweed (Hieracium) and Beans (Lablab). Detailed investigation by S. Blixt on pea plant led to locate Mendel's seven characters on 4 different chromosomes, i.e., 1, 4, 5 and 7.
However, Mendel's work did not receive any recognition, it deserved, till 1900.
Mendel's work remained unnoticed and unappreciated for several years due to following reasons:
(1) Communication was not easy in those days and his work could not be widely publicised.
(2) His concept of stable, unblending, discrete units or factors for various traits did not find acceptance from the contemporaries.
(3) His approach of using mathematical and statistical analysis to explain biological phenomena was totally new and unacceptable to many of the biologists of that time.
(4) He could not provide any physical proof for the existence of factors. It was rediscovery of his work by a Dutch -Hugo de Vries, a German -Carl Correns and an Austrian botanist -Erich von Tschermak, independently in 1900, that brought Mendel to limelight. Correns raised status of Mendel's generalisations to laws.
Selection of pea plant : The main reasons for adopting garden pea (Pisum sativum) for experiments by Mendel were as follows :
(1) Pea has many distinct alternative traits (clear contrasting characters).
(2) Life span of pea plant is short.
(3) Flowers show self (bud) pollination, so are true breeding.
(4) It is easy to artificially cross-pollinate the pea flowers. The hybrids thus produced were fertile.
Concept Builder
1. Term allele was given by Bateson, term homozygous and heterozygous were given by Bateson and Saunders; genotype, phenotype, gene and pureline by Johannsen.
2. Father of genetics -Mendel; Father of Modem genetics -Bateson
3. Isoalleles : Alleles that produce similar phenotypes but are distinguishable amongst themselves through changed optima, e.g.,  .
.
4. Pseudoalleles : Genes are present together side by side and they produce related phenotypes. These are distinguished from true alleles through rare crossing over, e.g., star (dominant) and asteroid (recessive) traits in Drosophila.
5. Pure lines (pure breeding line) : A population obtained by continuous inbreeding over many generations, such that each individual has essentially the same genome as every other member of the inbred line and that all (or most) loci are homozygous.
Mendel's Work and Results
Mendel has made cross between parents having contrasting traits.
Firstly he made monohybrid cross (cross between parents, differ from each other in one character) followed by dihybrid cross (cross between parents, differ from each other in two characters) and then trihybrid.
The F1 hybrids were self crossed to give rise to F2 generation.
Mendel also carried out the reciprocal crosses and found that reciprocal crosses gave the same result.
(Reciprocal cross means opposite cross, i.e., the parent which provides male gamete in one cross, in second experiment it provides the female gamete and vice versa).
The result of reciprocal cross proves that both gametes produce the same effect and it does not matter which parent provides male and which one provides female gamete.
On the basis of his experimental crosses, he formulated four postulates.
Postulate I:
According to this postulate characters are controlled by a pair of unit factors. The two factors are now called alleles or allelomorphic pair.
Postulate II :
If two dissimilar unit factors are present in an individual, only one expresses itself. The one which expresses itself is known as dominant factor, while the second which does not express at all is known as recessive factor.
Postulate III :
According to this postulate, two contrasting alleles responsible for contrasting traits present in an individual do not get mixed and get separated from each other at the time of gamete formation by F1 hybrid and due to their recombination, four combinations can be obtained in equal frequency.
All the above three postulates are based upon Mendel's monohybrid cross or one gene interaction.
Law of dominance and law of segregation can be explained on the basis of monohybrid cross or one gene interaction.
(1) Law of dominance :
This law states that when two contrasting alleles for a character come together in an organism, only one is expressed completely and shows visible effect.
It is called dominant and the other allele of the pair which does not express and remains hidden is called recessive.
This law is not universally applicable.
Plant height is controlled by two alleles -Dominant allele (T) and Recessive allele (t)
These two alleles can be present in three forms

Tt -Dominant allele expressed] Heterozygous (Hybrid for character)
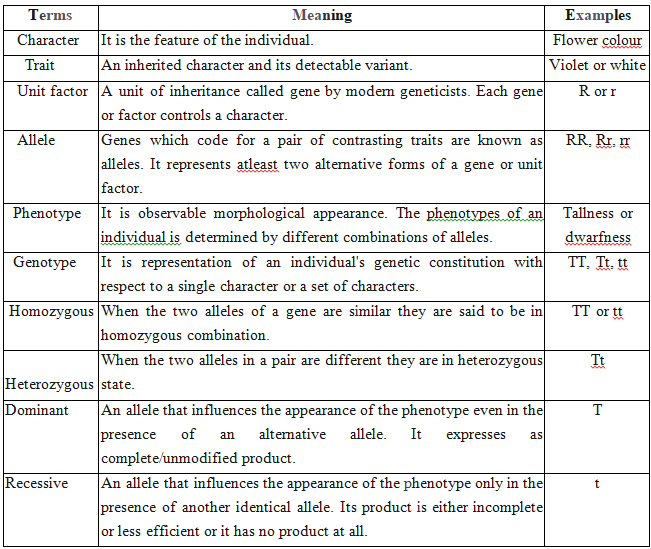
Mendel crossed two pea plants, one homozygous tall (TT) and another homozygous dwarf (tt).
He observed that all the F1 progeny plants were tall, like one of the parents, none were dwarf.
He made similar observations for the other pair of traits and found that F1 always resembled only one of the parents, and that the trait of other parent was not seen in them.
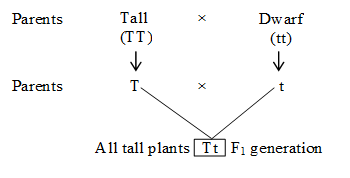
(2) Law of segregation or Law of purity of gametes:
This law states that both parental alleles (recessive and dominant) of F1 separate and are expressed phenotypically in F2 generation. This law is universally applicable.
The F2 generation was produced by allowing F1 hybrid to self pollinate, to find out segregation or separation.
It was observed that both dominant and recessive plants appeared in 3 : 1 ratio. Thus, F2 progeny shows both parental forms.
On the basis of F2 generation, following observations can be made:
(i) An organism generally has two alleles for each character. These alleles may either be similar or dissimilar. Organism with similar alleles of a pair is called pure or true breeding for that character. If the organism contains dissimilar alleles of a pair, the organism is impure or hybrid.
(ii) An organism receives one of the two alleles from the male gamete and the other from the female gamete. The gametes fuse during fertilization and form a zygote. Zygote develops into an organism.
(iii) Each gamete (male or female) has only one allele of the pair. Thus, each gamete is pure for a trait. That is why this law is often called as Law of purity of gametes.
(iv) The fusion between male and female gametes to produce zygote is a random process.
The plants obtained in F2 generation show 3 (tall) : 1 (dwarf) phenotypic ratio. Of these three tall plants, one is pure or homozygous dominant and the remaining two are heterozygous (tall in this case). There is only one plant that shows recessive character (dwarf in this case). Dwarf is pure or true breeding, being homozygous recessive.
Postulate IV :
This postulate was made on the basis of dihybrid cross or two genes interaction.
He postulated that inheritance of one character is independent of the inheritance of another character.
On the basis of this postulate, Mendel proposed the "Law of independent assortment".
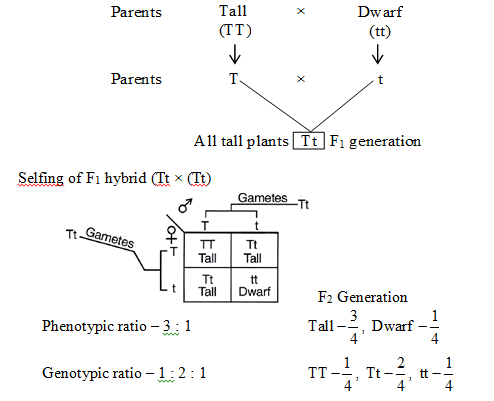
(3) Law of independent assortment :
The law of independent assortment states that when a cross is made between two individuals different from each other in two or more characters, then the inheritance of one character is independent of the inheritance of another character.
Because of their independent assortment, besides the parental types, recombinants are also obtained.
In dihybrid cross, these combinations are obtained in the ratio of 9 : 3 : 3 : 1. e.g., He crossed homozygous dominant round and yellow seeded plant (RRYY) with homozygous recessive wrinkled and green seeded (rryy) plant.
The F1 hybrids were all heterozygous, showing yellow and round seeded plants. This law is not universally applicable.
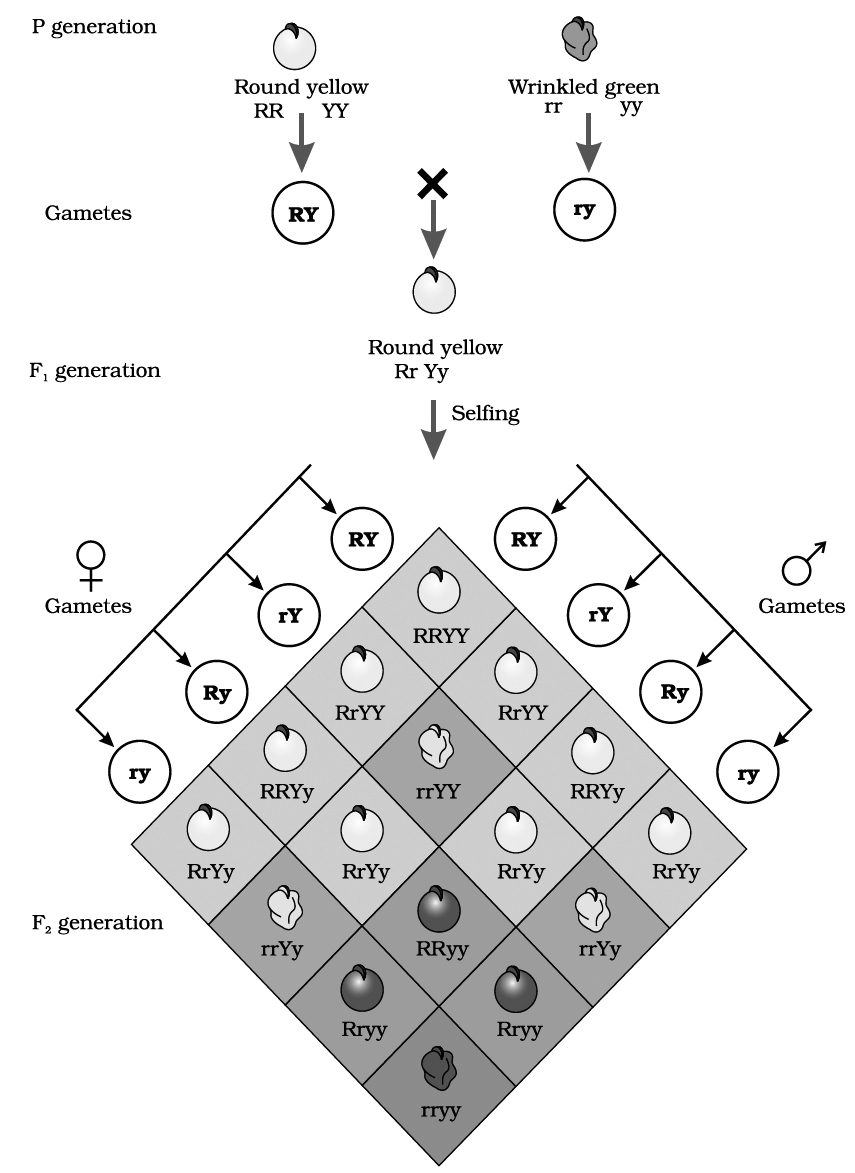
two pairs of contrasting traits : seed colour and seed shape
Phenotypic ratio: round yellow : round green : wrinkled yellow : wrinkled green
9/16 3/16 3/16 1/16
Genotypic ratio : 1 : 2 : 1 : 2 : 4 : 2 : 1 : 2 : 1
If the phenotypic ratio of each pair of alleles (e.g., yellow and green colour of seed) is considered, it shows 12(9 + 3) yellow seeded plants and 4(3 +1) green seeded plants.
This comes to 3 : 1 ratio; similar to one obtained in F2 generation of monohybrid cross showing segregation.
The same is true for another pair of alleles involved, i.e., round and wrinkled seeded plants. So, the results of each character are similar to the monohybrid ratio.
SUMMARISED ACCOUNT OF MENDEL'S EXPERIMENTS:
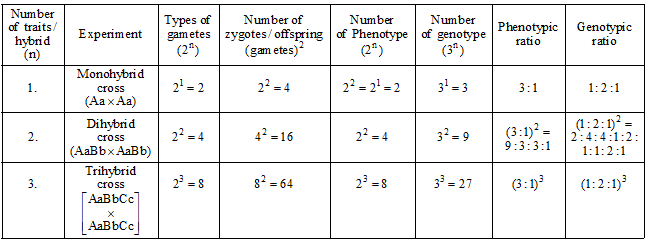

Back Cross and Test Cross
Back cross:
F1 hybrids are obtained by crossing two plants of parental generation.
Mendel devised a cross where F1 hybrid is crossed with anyone of the two parents, i.e., homozygous dominant and homozygous recessive.
Thus, there would be two possibilities:
(a) F1 hybrid (Tt) is crossed with homozygous dominant (TT)
(b) F1 hybrid (Tt) is crossed with homozygous recessive (tt)
Both these crosses collectively are called as back cross. If F1 is crossed with dominant parent, it is called out cross.
Test Cross:
Out of the two types of back crosses, a cross between F1 hybrid (Tt) and its homozygous recessive parent (tt) is called test cross.
This cross is called test cross because it helps to find out whether the given dominant F1 phenotype is homozygous or heterozygous.
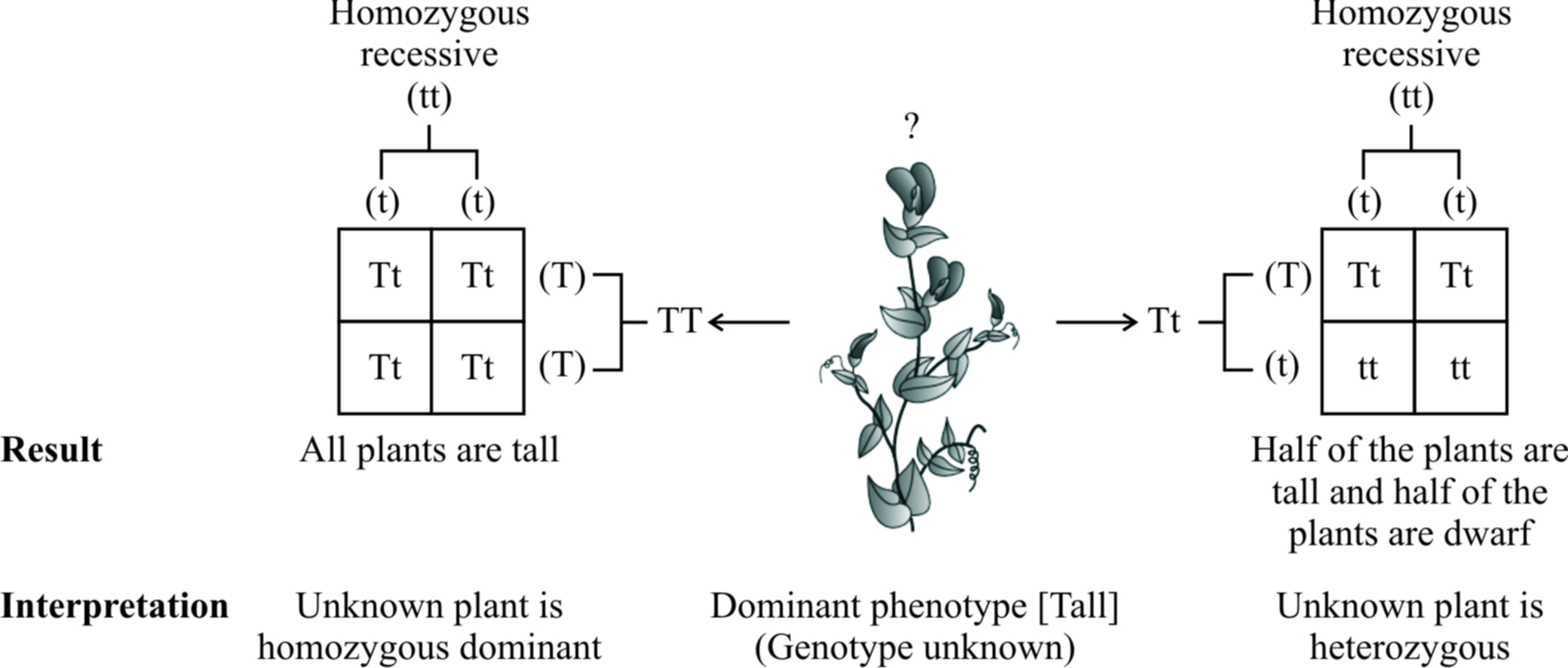
A monohybrid test cross between F1 tall plant (Tt) and its homozygous recessive parent (tt) will produce 50% heterozygous tall (Tt) and 50% homozygous recessive (tt), i.e., 1 : 1 ratio, for both phenotype and genotype.
If a test cross with two characters, i.e., dihybrid test cross is made, it gives four types of plants in 1 : 1 : 1 : 1 ratio.
The phenotypes obtained are similar to those found in F2 generation of dihybrid cross.
Thus, a dihybrid test cross between F1 yellow and round seeded plant (YyRr) and its homozygous recessive parent green and wrinkled (yyrr) would give following combinations:
1 yellow, round (YyRr) Parental combination 25%
1 yellow, wrinkled (Yyrr) Recombinants 25%
1 green, round (yyRr) Recombinants 25%
1 green, wrinkled (yyrr) Parental combination 25%
If this ratio is obtained, it would be confirmed that F1 hybrid with dominant phenotype is infact heterozygous.
The parental combinations (50%) are equal to the frequency of recombinants (50%).
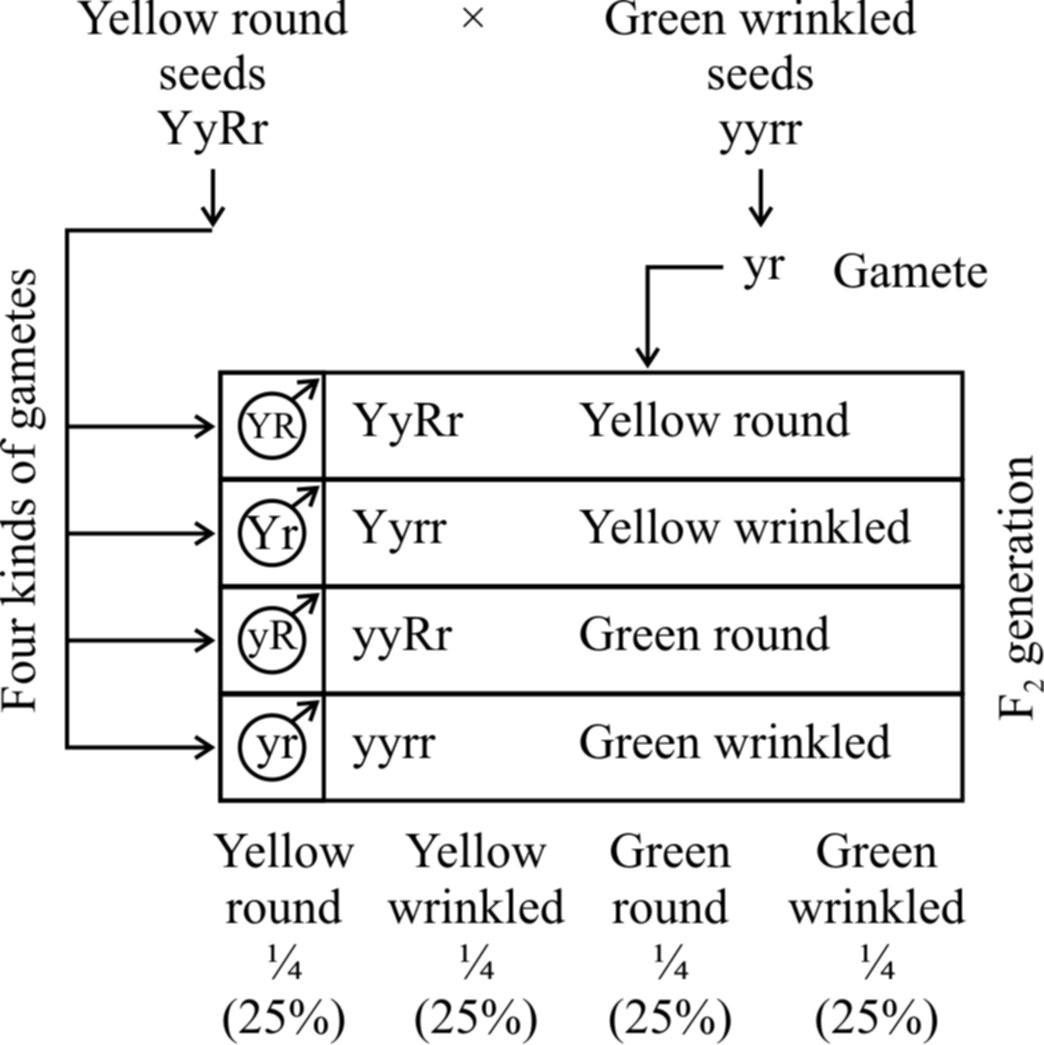
Trihybrid cross.
Mendel crossed two pea plants which differed in 3 characters and observed independent assortment of genes in them.
He crossed two pea plants pure in three traits viz., height of stem, form of seed and colour of cotyledon of seed.
He crossed homozygous tall, round and yellow (TT RR YY) plant with dwarf, wrinkled and green (tt rr yy).
All the F1 individuals produced were tall, round and yellow (Tt Rr Yy) and are called trihybrids.
On selfing trihybrids, F2 phenotypic ratio is 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1. The ratio for a trihybrid test cross is 1 : 1 : 1 : 1 : 1 : 1 : 1 : 1 .
Inheritance of one gene
- Books Name
- A TEXT OF BIOLOGY - CLASS XII
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 12
- Subject
- Biology
ONE GENE INTERACTION (w.r.t. Post-Mendelian inheritance)
(1) Incomplete dominance:
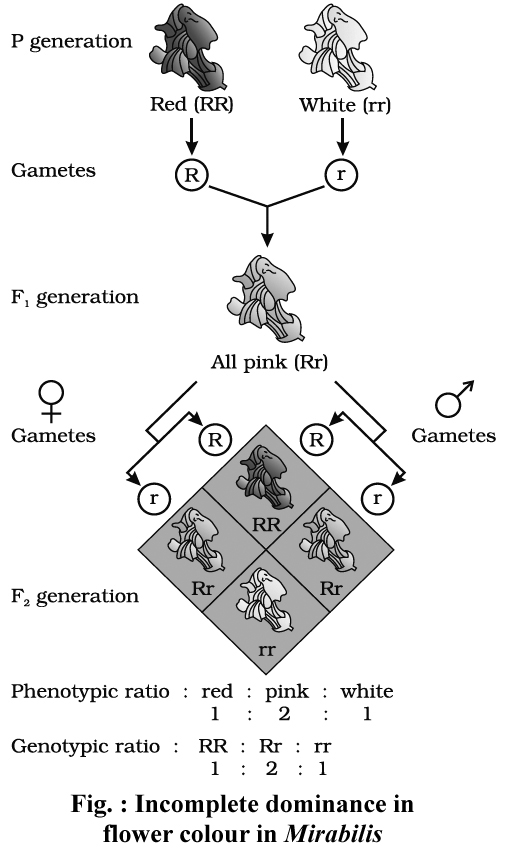
After Mendelism a few cases were observed where F1 phenotype is intermediate between dominant and recessive phenotypes.
The most common example of incomplete dominance is that of flower colour in Mirabilis jalapa (Gulbansi or 4'O clock plant), studied by Carl Correns.
Homozygous red (RR) flowered variety was crossed with white (rr) flowered variety.
F1 offspring had pink flowers.
Thus, here one allele is incompletely dominant over the other so that intermediate phenotype is produced by F1 hybrid with respect to the parents.
This is called incomplete dominance.
Incomplete dominance for flower colour [Red(RR), Pink(Rr), White (rr)] is also known to occur in Antirrhinum majus (Snapdragon or Dog flower).
The phenotypic ratio and genotypic ratio in F2 generation is identical in case of incomplete dominance i.e., 1 : 2 : 1.
Explanation of the concept of dominance:
Every gene contains information to express a particular trait.
Diploid organisms have two copies of each gene, they are called alleles.
These two alleles may be identical or non-identical.
One of them may be different due to some changes that it has undergone which modifies the information that particular allele contains.
Theoretically, the modified allele could be responsible for production of
(i) The normal/less efficient enzyme, or
(ii) a non-functional enzyme, or
(iii) no enzyme at all
In case (i), the modified allele is equivalent to the unmodified allele, i.e., it will produce the same phenotype/trait.
But, if the allele produces a non-functional enzyme or no enzyme [case (ii) &(iii)], the phenotype may be effected.
The unmodified (functioning) allele, which represents the original phenotype is the dominant allele and the modified allele is generally the recessive allele.
Hence, the recessive trait is due to nonfunctional enzyme or because no enzyme is produced.
If the mutated allele forms an altered but functional product, it behaves as incompletely or codominant allele.
(2) Multiple allelism:
Mendel proposed that each gene has two contrasting forms, i.e., alleles.
But there are some genes which are having more than two alternative forms (allele).
Presence of more than two alleles for a gene is known as multiple allelism.
Multiple alleles are present on the same locus of homologous chromosome.
Multiple alleles can be detected only in a population.
A well known example to explain multiple alleles in human beings is ABO blood type.
Landsteiner discovered ABO system of blood groups. The fourth group AB was discovered by de Castello and Steini.
Bernstein showed that these groups are controlled by 3 alleles -IA, IB and IO/i.
These alleles are autosomal and follow Mendelian pattern of inheritance.
The alleles IA and IB produce a slightly different form of the sugar while IO doesn't produce any sugar. Because humans are diploid organism, each person possesses any two of the three I gene alleles.
IA and IB are completely dominant over IO, but when IA and IB are present together they both express their own types of sugar thus, behaving as codominant alleles.
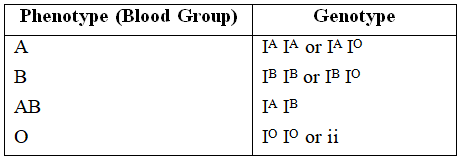
Possible blood types of children from parents of various blood types.

Other examples of multiple alleles are - coat colour in rabbit, eye colour in Drosophila and self incompatibility in tobacco. Formula to find out number of genotypes for multiple allelism is n/2(n +1) , where 'n' is number of alleles.
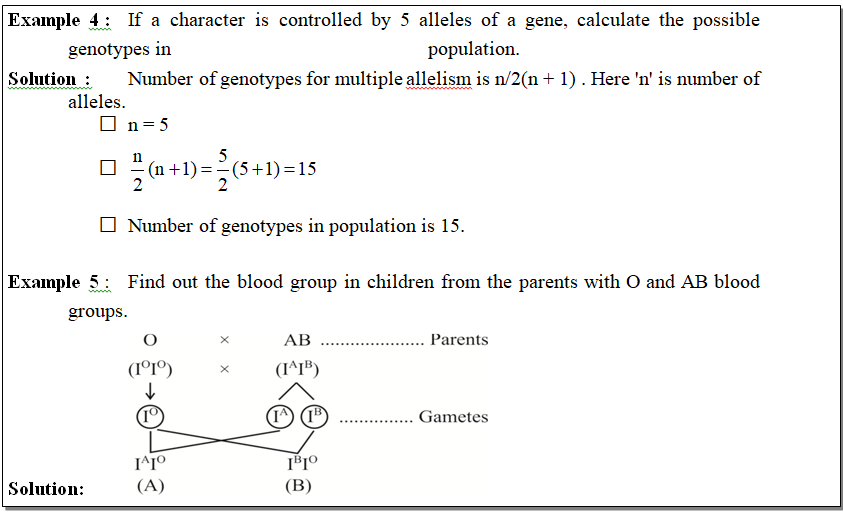
The possibility of blood groups in offsprings of this couple is A and B.
(3) Co-dominance.
In co-dominance, the genes of an allelomorphic pair are not related as dominant and recessive but both of them express themselves equally in F1 hybrids.
These follow the law of segregation and F2 progeny exhibits 1 : 2 : 1 ratio. Heterozygous for sickle cell anaemia (HbAHbS), AB and MN blood groups are examples of co-dominance of alleles.
(4) Lethal genes or lethality.
A lethal gene usually results in the death of an individual when present in homozygous condition. Most striking example to explain lethal gene is sickle cell anaemia (HbSHbS).
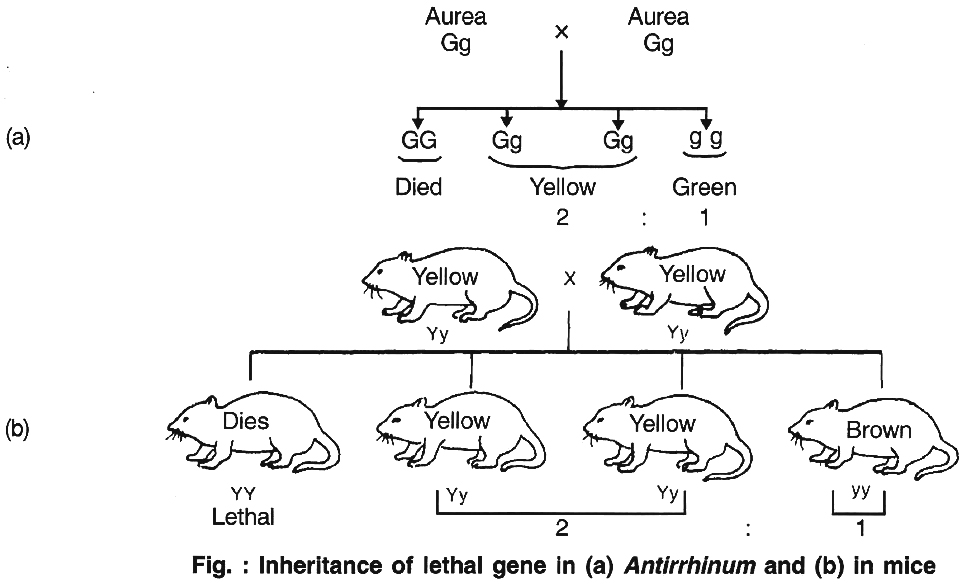
Cuenot (1905) first reported that inheritance in mouse body colour did not agree with Mendelian inheritance, because the dominant allele for yellow body colour is lethal in homozygous condition.
The homozygous dominant gene carrying mouse died, proving dominant gene is lethal in homozygous form.
This is called absolute lethality. In plants, it was first reported in Antirrhinum majus by E. Baur, where yellow leaved or golden leaved or Aurea plant never breed true. Thus, the ratio comes out to be 2 : 1.
(5) Pleiotropic genes:
The ability of a gene to have multiple phenotypic effects (as it influences a number of characters simultaneously) is known as pleiotropy.
The gene having multiple phenotypic effects is called pleiotropic gene.
It is not essential that the traits are equally influenced. Sometimes, the effect of the gene is more evident in case of one trait (major effect) and less evident in case of others (secondary effect).
Occasionally, a number of related changes are caused by a gene.
They are together called syndrome.
Some common examples in humans are -Cystic fibrosis, Marfan syndrome and Phenylketonuria, while in Drosophila, a single gene influences the size of wings, character of balancers, position of dorsal bristles, eye colour, shape of spermathecae, fertility and longevity.
In human beings pleiotropy is exhibited by sickle cell anaemia in heterozygous condition (HbAHbS).
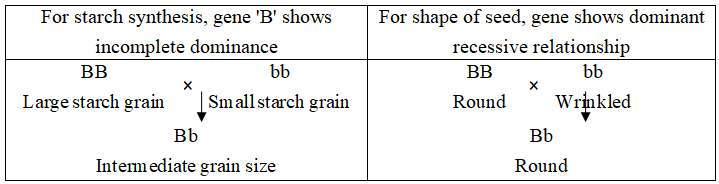
In case of pea, the gene which controls the starch synthesis also controls the shape of the seed.
Concept Builder
(1) Rh factor: It is an antigen protein on RBC surface, reported by Landsteiner and Wiener in blood of Indian brown Rhesus monkey (Macacus rhesus). 93% Indians have this antigen and are called Rh positive (Rh+).
(2) Marian syndrome: Slender body, elongation of limbs, hypermobility of joints, dislocation of lens and susceptibility to heart diseases.
Inheritance of two genes
- Books Name
- A TEXT OF BIOLOGY - CLASS XII
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 12
- Subject
- Biology
TWO GENES INTERACTION (w.r.t. Post-Mendelism)
Genes usually function or express themselves singly or individually.
But, many cases are known where two genes of the same allelic pair or genes of two or more different allelic pairs influence one another.
This is called gene interaction.
Non-allelic genetic interactions.
These are interactions between genes located on the same chromosome or on different but non-homologous chromosomes controlling a single phenotype to produce a different expression.
Each interaction is typical in itself and ratios obtained are different from those of the Mendelian dihybrid ratios.
Some of these interactions of genes are explained here which fall under this category and deviate from Mendel's ratios.
1. Complementary genes :
The complementary genes are two genes present on separate loci that interact together to produce dominant phenotypic character, neither of them if present alone, can express itself. It means that these genes are complementary to each other.
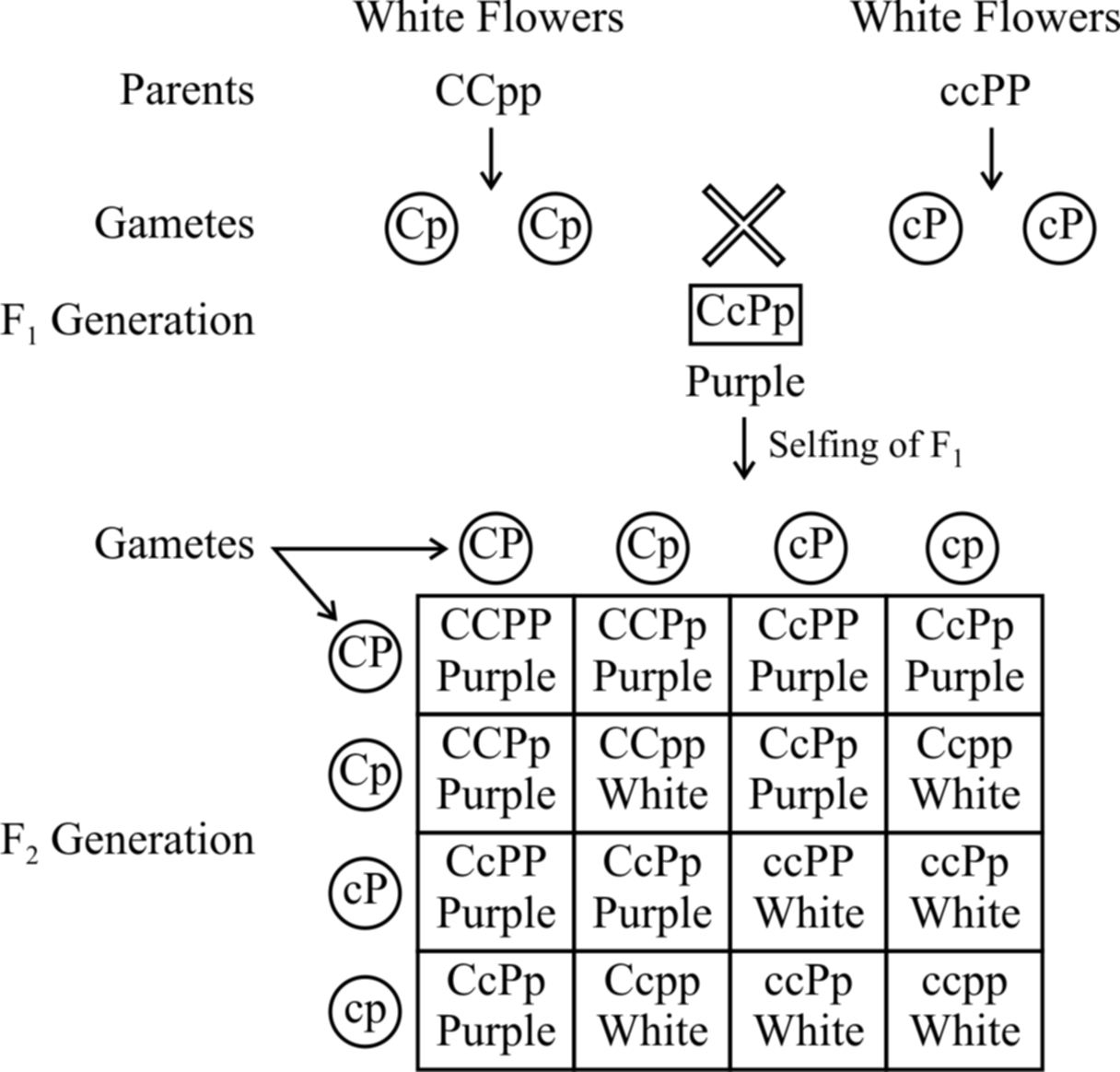
Fig. : Results of an experiment showing inheritance of flower colour in
Lathyrus odoratus controlled by complementary genes
Bateson and Punnet have demonstrated that in sweet pea (Lathyrus odoratus) purple colour of flowers develop as a result of interaction of two dominant genes C and P.
In the absence of dominant gene C or P or both, the flowers are white.
It is believed that gene C produces an enzyme that catalyzes the formation of necessary raw material for the synthesis of pigment anthocyanin and gene P produces an enzyme which transforms the raw material into the pigment.
It means the pigment anthocyanin is the product of two biochemical reactions, the end product of one reaction forms the substrate for the other.
![]()
Therefore, if a plant has ccPP, ccPp, CCpp or Ccpp genotypes, it bears only white flowers. Purple flowers are formed in plants having genotype CCPP or CCPp or CcPP or CcPp.
From checker board, it is clear that 9 : 7 ratio between purple and white is a modification of 9 : 3 : 3 : 1 ratio.
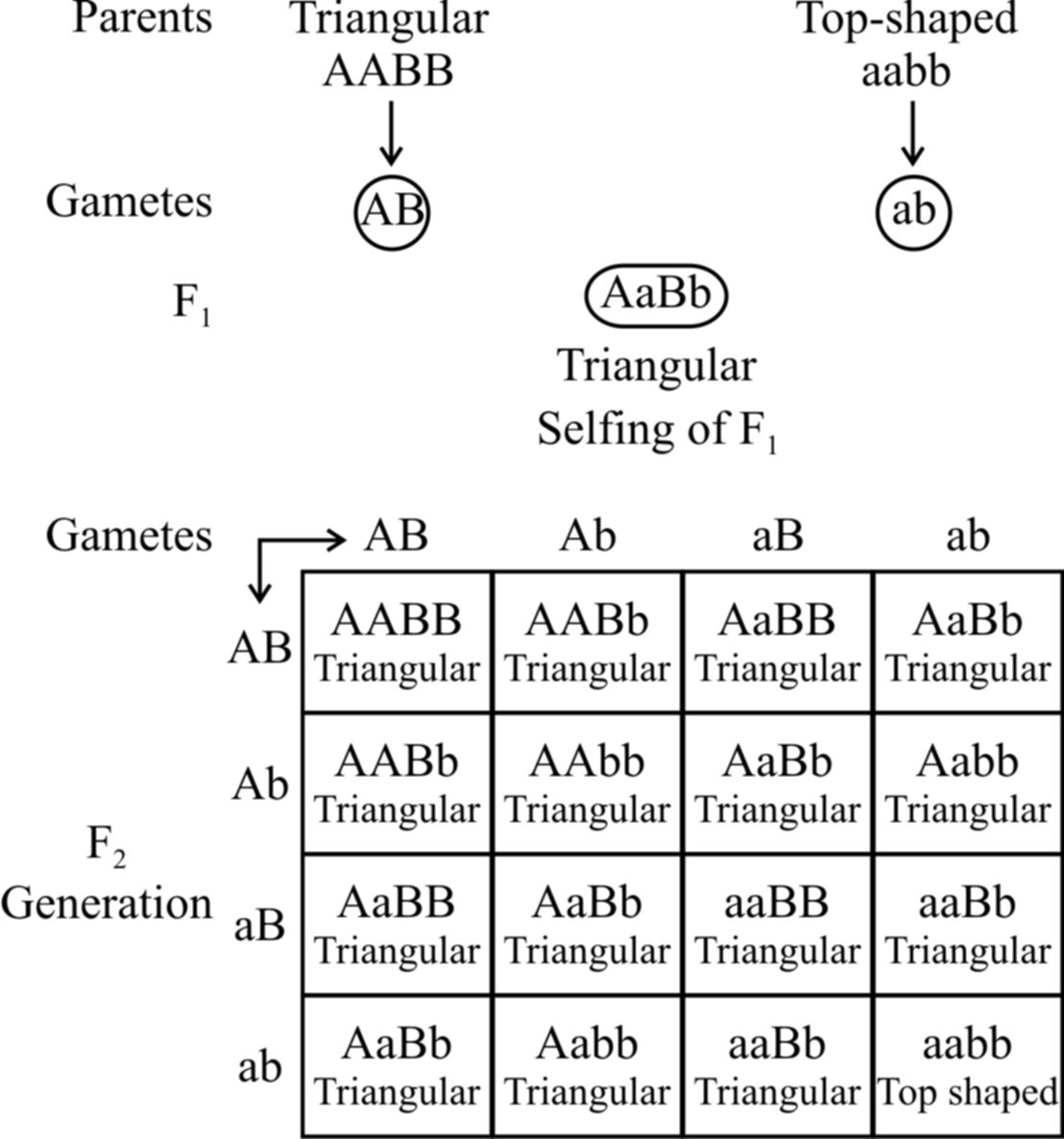
2. Duplicate genes.
If the dominant alleles of two gene loci produce the same phenotype, whether inherited together or separately, the 9 : 3 : 3 : 1 ratio is modified into a 15 : 1 ratio.
Example: The capsules of shepherd's purse (Capsella) occur in two different shapes, i.e., triangular and top-shaped. When a plant with triangular capsule is crossed with one having top-shaped capsule, in F1 only triangular character appears. The F1 offspring by self crossing produced the F2 generation with the triangular and top-shaped capsules in the ratio of 15 : 1. Two independently segregating dominant genes (A and B) have been found to influence the shape of capsule in the same way. All genotypes having dominant alleles of both or either of these genes (A and B) would produce plants with triangular-shaped capsules. Only those with the genotype aabb would produce plants with top shaped capsules. The results of this example are given below.
3. Epistasis
A gene which masks (hides) the action of another gene (non allelic) is termed as epistatic gene. The process is called epistasis. The gene whose effects are masked is called hypostatic gene.
Epistasis is of two types :
(a) Recessive epistasis :
Here the recessive allele in homozygous condition masks the effect of dominant allele, e.g., in mice, the wild body colour is known as agouti (greyish) and is controlled by a gene say A which is hypostatic to recessive allele c.
The dominant allele C in the presence of 'a' gives coloured mice.
In the presence of dominant allele C, A gives rise to agouti.
So, CCaa will be coloured and ccAA will be albino.
When coloured mice (CCaa) are crossed with albino (ccAA), agouti mice (CcAa) appear in F1.
cc masks the effect of AA and is therefore epistatic.
Consequently, cc AA is albino.
The ratio 9 : 3 : 3 : 1 is modified to 9 : 3 : 4.
The combination ccaa is also albino due to the absence of both the dominant alleles.
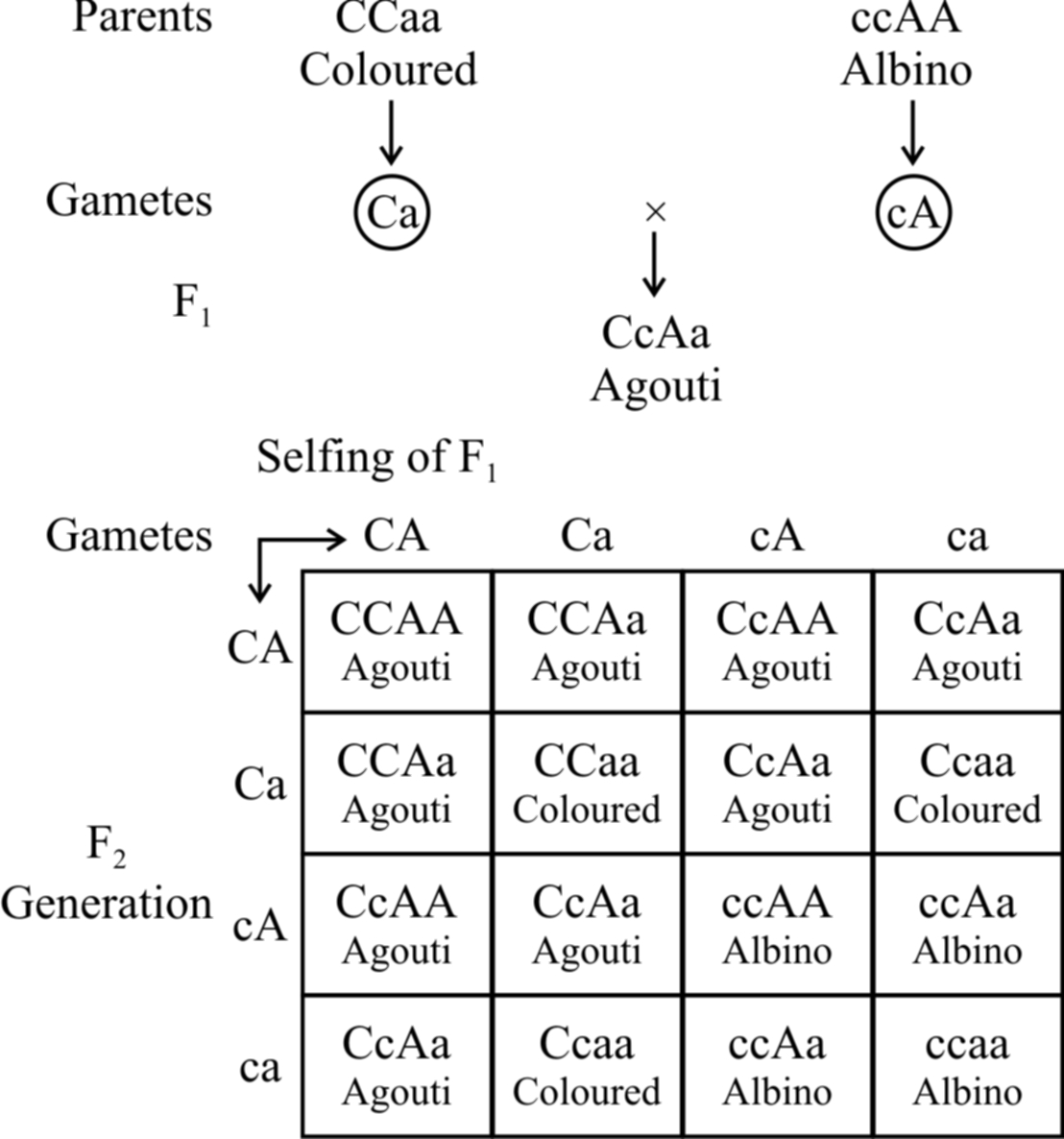
Fig. : Recessive epistasis
(b) Dominant epistasis :
In summer squash or Cucurbita pepo, there are three types of fruit colour -yellow, green and white.
White colour is dominant over other colours, while yellow is dominant over green.
Gene for white colour (W) masks the effects of yellow colour gene (Y).
So, yellow colour is formed only when the dominant epistatic gene is represented by its recessive allele (w). When the hypostatic gene is also recessive (y) , the colour of the fruit is green.
White Fruit – W – Y –, W – y –
Yellow Fruit – wwY –
Green Fruit – wwyy
A cross between a pure breeding white summer squash, (WWYY) with a pure breeding green summer squash, (wwyy) yields white fruits in the F1 generation. Upon selling of F1, the F2 generation comes to have 12 white fruit : 3 yellow fruit : 1 green fruit.
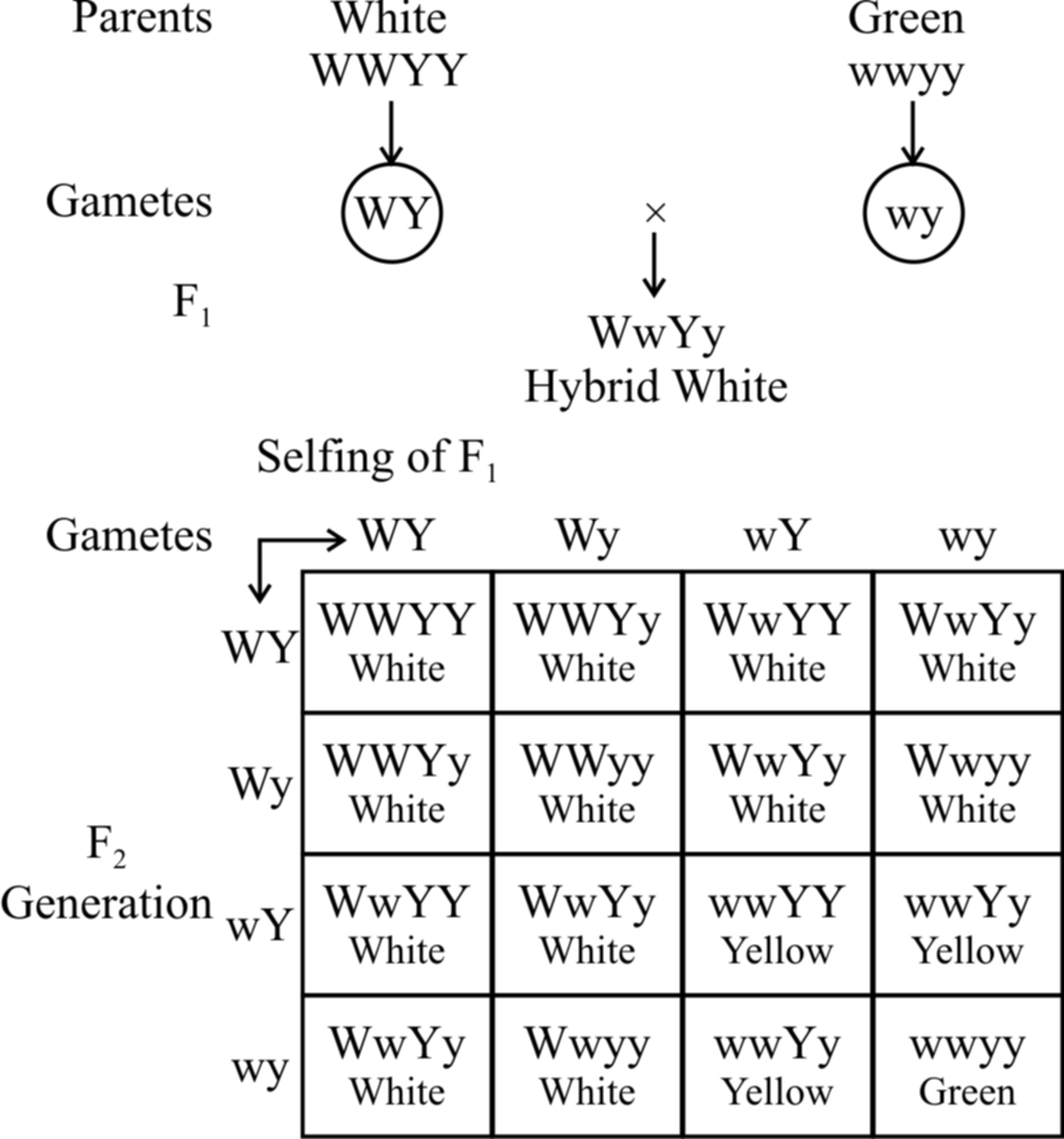
Fig. : Dominant epistasis
Polygenic inheritance or quantitative inheritance
Quantitative inheritance is controlled by two or more genes in which the dominant alleles have cumulative effect, with each dominant allele expressing a part of functional polypeptide and full trait is shown when all the dominant alleles are present.
Genes involved in quantitative inheritance are called polygenes.
H. Nilsson-Ehle (1908) and East (1910) demonstrated segregation and assortment of genes controlling quantitative traits, e.g., Kernel colour in wheat and corolla length in tobacco.
Kernel colour in wheat. Swedish geneticist, H. Nilsson-Ehle (1908) crossed red kerneled variety with white kerneled variety of wheat.
Grains of F1 were uniformly red but intermediate between the red and white of parental generation.
When members of F1 were self·crossed among themselves, five different phenotypic classes appeared in F2 showing the ratio of 1 : 4 : 6 : 4 : 1.
(i) The extreme red -1/16 (as red as to the parent of F1)
(ii) Deep red (Dark red) -4/16
(iii) Intermediate red -6/16 (similar to F1)
(iv) Light red -4/16
(v) White -1/16 (as white as to the parent of F1)
Nilsson Ehle found that the kernel colour in wheat is determined by two pairs of genes AA and BB.
Gene A and B determine the red colour of kernel and are dominant over their recessive alleles. Each gene pair shows Mendelian segregation.
Heterozygotes for two pairs of genes (AaBb) segregate into 15 red and one white kerneled plants.
But all the red kernels do not exhibit the same shade of redness.
The degree of redness was found to correspond with the number of dominant alleles.
Skin Colour in Man
The presence of melanin pigment in the skin determines the skin colour.
The amount of melanin developing in the individual is determined by three (two also) pairs of genes.
These genes are present at three different loci and each dominant gene is responsible for the synthesis of fixed amount of melanin.
The effect of all the genes is additive and the amount of melanin produced is always proportional to the number of dominant genes.
Subsequent studies after Davenport have shown that as many as six genes may be involved in controlling the skin colour in human beings.
As shown in table, the effect of all the genes is additive (The character is assumed to be fixed by three pairs of polygenes).
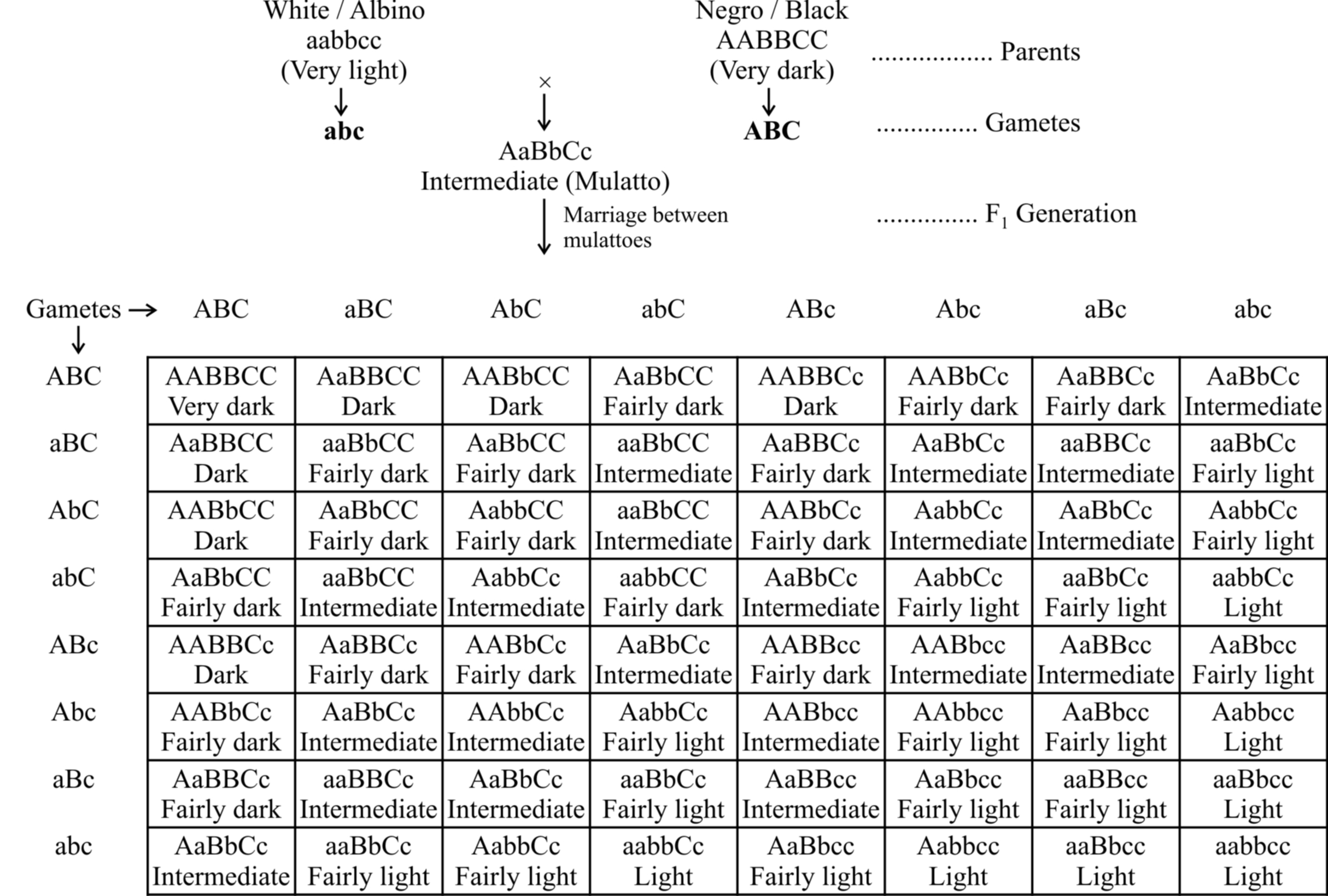
Phenotypes: 1 (Very dark) : 6 (Dark) : 15 (Fairly dark) : 20 (Intermediate) : 15 (Fairly light) : 6 (Light) : 1 (Very light)
The F1 progeny between an albino and a negro individual called mulatto produces intermediate skin colour.
In F2 generation, the coloured offsprings exhibit different shades in the ratio 1 : 6 : 15 : 20 : 15 : 6 : 1.
The frequency distribution for skin colour can be represented either as a histogram or in the form of a bell-shaped normal distribution curve.
Looking at the histogram, it can be concluded that in polygenic inheritance, the extreme phenotypes are rare and the intermediate ones are more frequent.
Some other example of quantitative traits are cob length in maize, human intelligence, milk and meat production, height in human and size, shape and number of seeds and fruits in plants.
- Number of phenotype for polygenes = 2n + 1
- Number of genotype for polygenes = 3n, where n represent pair of polygenes.
Example.7 : Calculate the sum total of phenotypes and genotypes in F2 generation if a character is controlled by 2 pair of polygenes.
Solution : Number of phenotype for polygenes = 2n + 1
(2 × 2 + 1) = 5
Number of genotype for polygenes = 3n
32 = 9
i.e., 5 + 9 =14
CHROMOSOMAL THEORY OF INHERITANCE / PARALLELISM BETWEEN
CHROMOSOMES AND MENDELIAN FACTORS
Chromosomal theory of inheritance was proposed independently by Sutton and Boveri.
The two workers found a close similarity between the transmission of hereditary traits and behaviour of chromosomes while passing from one generation to the next through the agency of gametes.
Sutton and Boveri noted that the behaviour of chromosomes is parallel to the behaviour of Mendelian factors (Genes).
The salient features of chromosomal theory of inheritance are as follows :
1. Like the hereditary traits the chromosomes retain their number, structure and individuality throughout the life of an organism and from generation to generation. The two neither get lost nor mixed up. They behave as units.
2. Both chromosomes as well as genes occur in pairs in the somatic or diploid cells. The two alleles of a gene pair are located on homologous sites on homologous chromosomes.
3. A gamete contains only one chromosome of a type and only one of the two alleles of a trait.
4. The paired condition of both chromosomes as well as Mendelian factors is restored during fertilization.
Homologous chromosomes synapse during meiosis and then separate or segregate independently into different cells which establishes the quantitative basis for segregation and independent assortment of hereditary factors.
Sutton united the knowledge of chromosomal segregation with Mendelian principles and called it the chromosomal theory of inheritance.
Johannsen (1909) coined the term gene, for mendelian factor.
Following the synthesis of ideas, experimental verification of the chromosomal theory of inheritance by T.H. Morgan and his colleagues, led to discovery of the basis for the variations, that sexual reproduction produced.
Thomas Hunt Morgan (1866-1945) is known as father of experimental genetics. He was awarded Nobel Prize of physiology in 1933 for his pioneer work in experimental genetics.
Drosophila melanogaster as material for experimental Genetics
Fruit fly Drosophila is a tiny fly of about 2 mm size which is found over ripe fruits like mango and banana.
The fly is actually attracted to yeast cells present on the surface of the ripe fruits. Drosophila is more suitable than pea as experimental material because of following reasons :
(i) It can be easily reared and bred under laboratory conditions.
(ii) The fly has a short life span of about two weeks. The fruit fly can be bred throughout the year so that numerous generations can be obtained in a single year instead of one as in case of Pea.
(iii) A single mating produces hundreds of offsprings.
(iv) Females are easily distinguishable from the males by the larger body size and presence of ovipositor (egg laying structure).
(v) The animal shows a number of externally visible and easily identifiable contrasting traits.
(vi) It has a smaller number (4 pairs) of morphologically distinct chromosomes.
(vii) Polytene chromosomes occur in the salivary glands of larva. Polytene chromosomes can be used to study different types of chromosome aberrations.
(viii) It has heteromorphic (XY) sex chromosomes in the male. The transmission of heteromorphic chromosomes can be easily studied from one generation to another.
LINKAGE (Exception to Law of Independent Assortment)
According to Mendel's law of independent assortment, the gene controlling different characters get assorted independent to each other.
It is correct if the genes are present on two different chromosomes, but if these genes are present on same chromosome they may or may not show independent assortment.
If crossing over takes place between these two genes then the genes get segregated and they will assort independent to each other. But if there is no crossing over between these two genes there is no segregation, hence only parental combination will be found in gametes.
The tendency of some of the genes to inherit together (in block) is known as linkage.
In 1906, Bateson and Punnet crossed two varieties of Lathyrus odoratus (sweet pea) and observed that the results do not agree with the Mendel's law of independent assortment.
They formulated the hypothesis of coupling and repulsion to explain the unexpected F2 results of dihybrid cross between a homozygous sweet pea having dominant alleles for blue flowers (BB) and long pollen grains (LL) with another homozygous double recessive plant with red flowers and round pollen grains (bbll).
Test cross ratio of 7 : 1 : 1 : 7 indicated that there was a tendency of the dominant alleles to remain together. Similar was the case with recessive alleles.
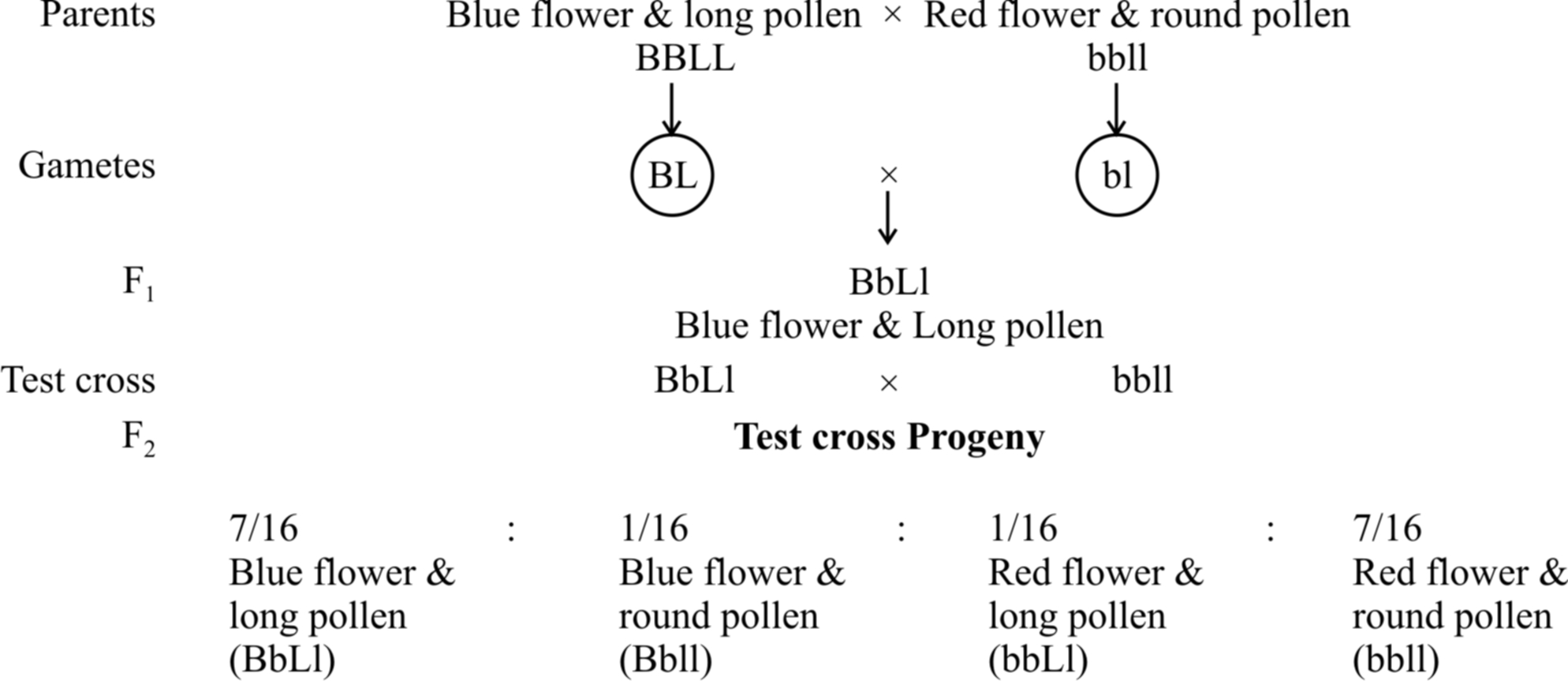
It was called gametic coupling by Bateson and Punnet.
Two dominant genes from one parent entered the same zygote more frequently than expected.
The tendency of two dominant genes to remain together in the process of inheritance was called as coupling.
In another cross, they took a sweet pea plant with blue flowers and round pollens (BBII) and other plant with red flowers and long pollens (bbLL) and obtained the ratio of 1 : 7 : 7 : 1 by test crossing F1 generation.
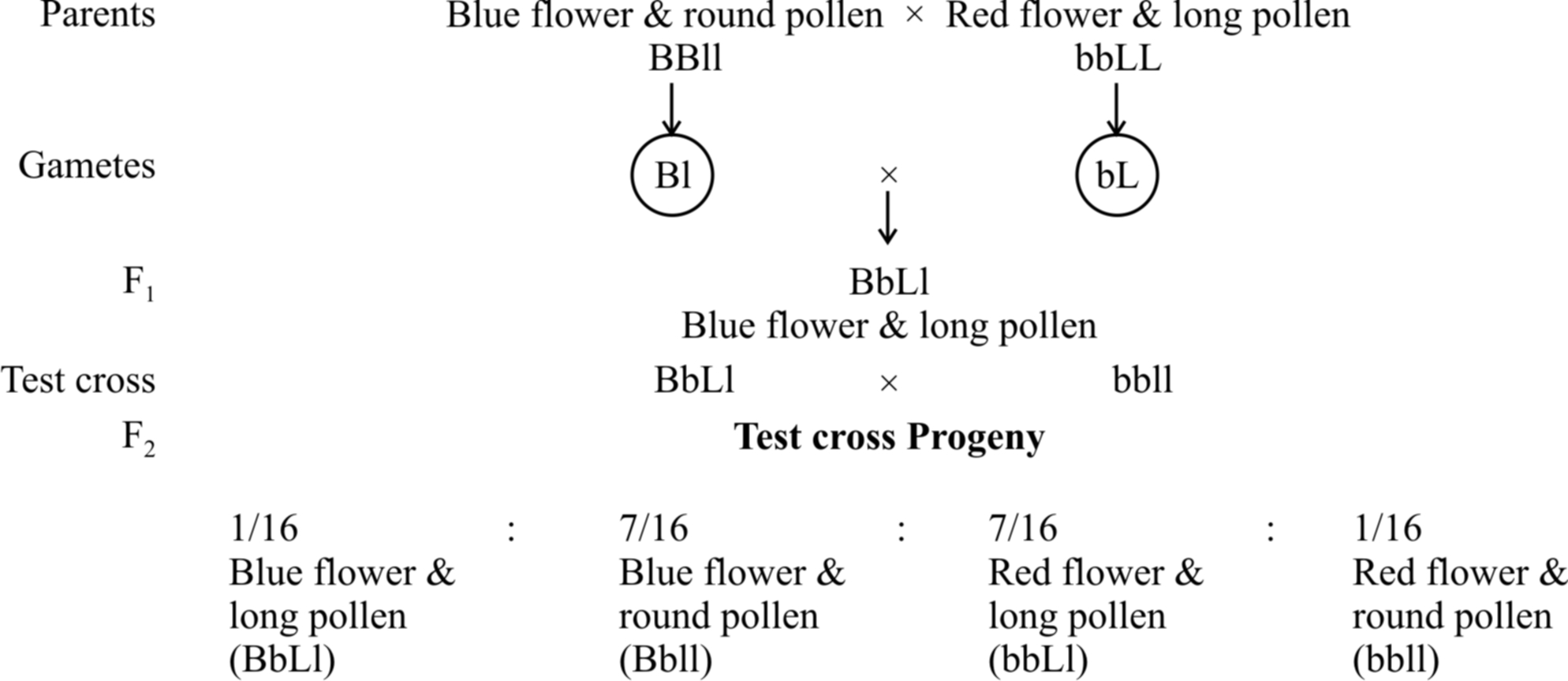
When two dominant or recessive genes come from different parents, they tend to remain separate hence, this ratio was called repulsion ratio.
T.H. Morgan in 1910 showed that coupling and repulsion are two aspects of the same phenomenon called linkage.
He suggested that the two genes present on the same chromosome, are in coupling phase and when present on two different homologous chromosomes are in repulsion phase.
Morgan carried out several dihybrid crosses in Drosophila to study genes that were sex-linked and
(A) At first, he crossed yellow bodied (y) and white eyed (w) female with brown bodied (y+) red eyed (w+) male which produced F1 with brown bodied red eyed female and yellow bodied white eyed male. In F2 generation, obtained by intercrossing of F1 hybrids, the ratio deviated significantly from expected. He found 98.7% to be parental and 1.3% as recombinants.
(B) In a second cross between white eyed and miniature winged female (wwmm) with wild red eyed (w+) normal winged male (m+), the F1 generation included red eyed normal winged female and white eyed miniature winged male. After intercrossing the F1 progeny, he found 62.8% parental and 37.2% recombinants.
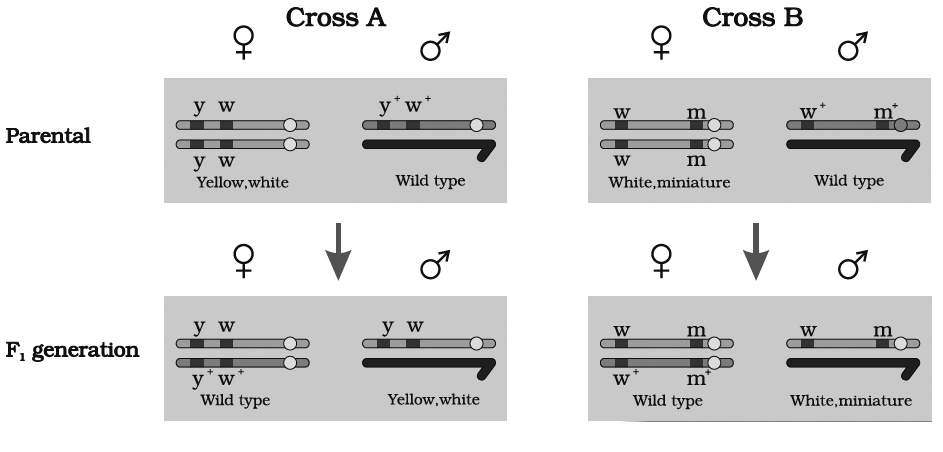
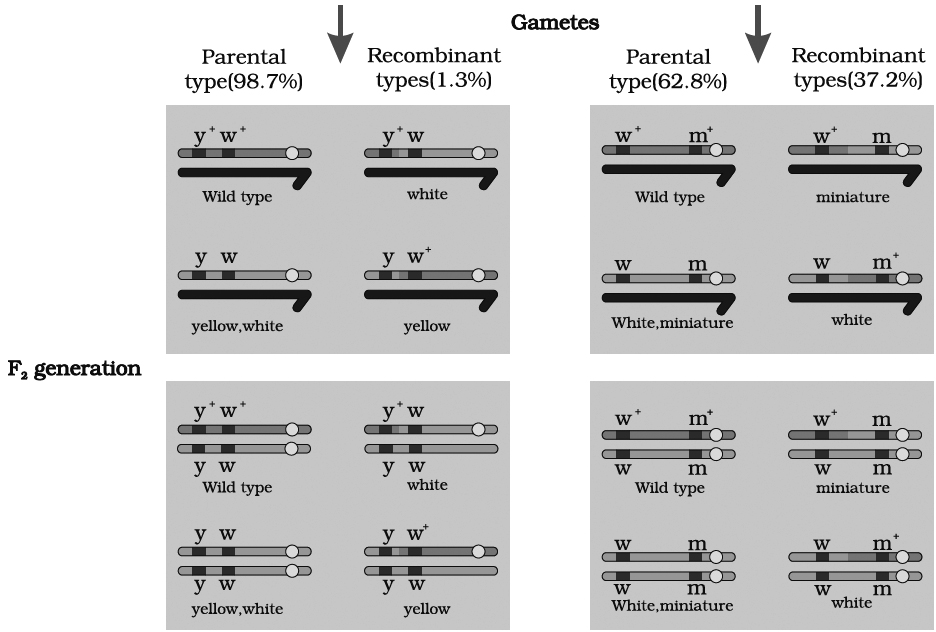
we can say that strength of linkage between y and w is higher than w and m
According to Morgan, the degree or the strength of linkage depends upon the distance between the linked genes in the chromosome.
Linkage, therefore, may be defined as "The tendency of two genes of the same chromosome to remain together in the process of inheritance".
Kinds of Linkage
T.H. Morgan and his coworkers found two types of linkage :
(1) Complete linkage.
Linkage of genes on a chromosome which is not altered and is inherited as such from generation to generation without any cross over.
In this type of linkage, genes are closely associated and tend to remain together.
Example: Male Drosophila and female silk worm (Bombax mort).
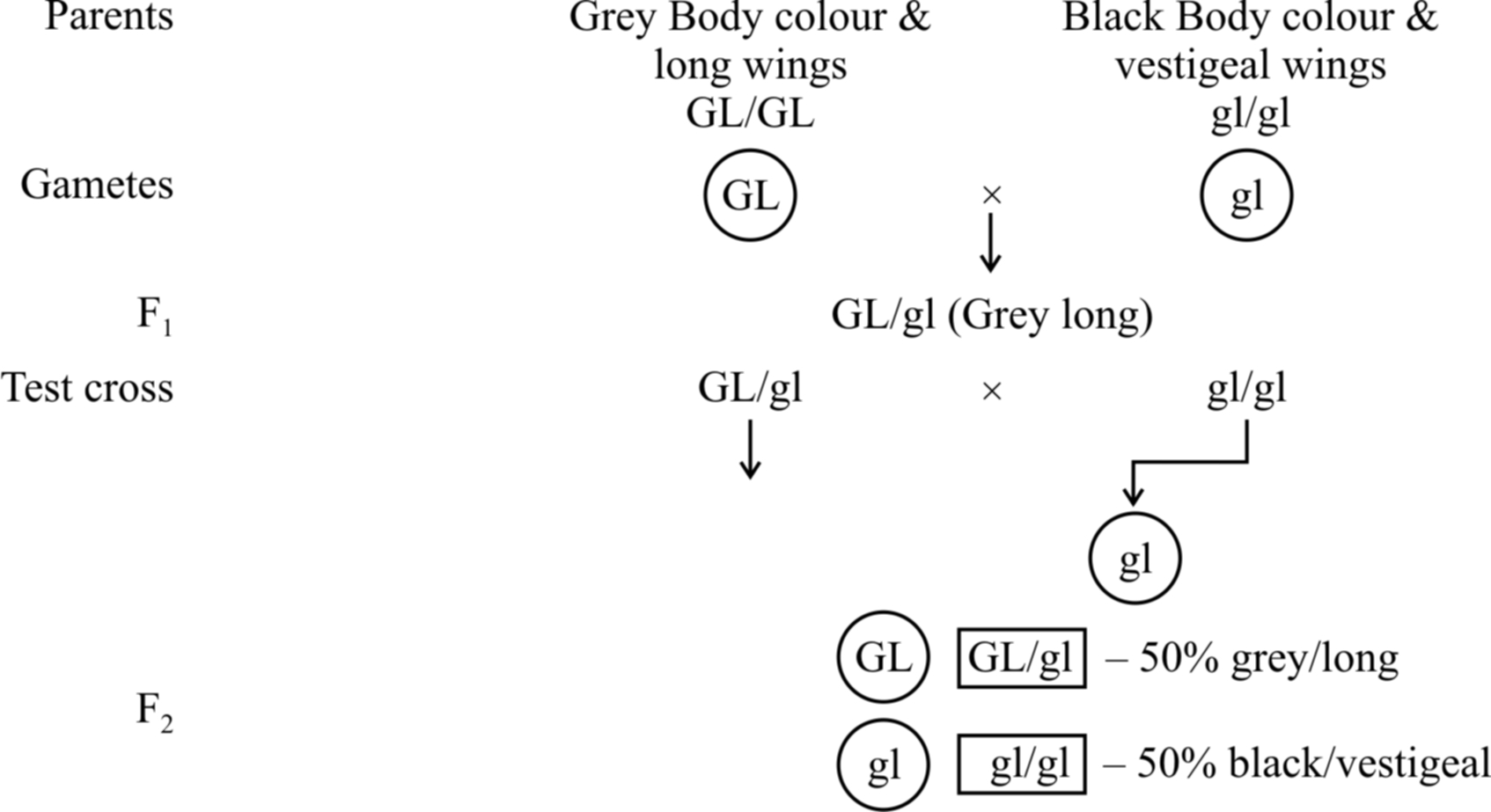
100% parental combinations indicated that the gene for grey body colour is completely linked with long wings.
In this dihybrid, F2 phenotypic ratio is 3: 1 and test cross ratio is 1 : 1 (like a monohybrid). Another example is inheritance of red eye and normal wing (PV/PV) with purple eye and vestigial wing character (pv/pv).
(2) Incomplete linkage.
The linked genes do not always stay together because homologous non sister chromatids may exchange segments of varying length with one another during meiosis.
This is known as crossing over.
The linked genes may have chances of separation by crossing over, are called incompletely linked genes and the phenomenon of their inheritance is called incomplete linkage.
It produces both parental and recombinant types in variable ratio.
Bateson and Punnet studied Lathyrus odoratus and defined coupling and repulsion of dominant and recessive gene.
In cis arrangement or coupling condition, the incomplete linkage ratio was 7 : 1 : 1 : 7 (14-parental 2-recombinants).
In trans arrangement or repulsion case, the ratio was 1:7:7:1 (parental 14, recombinants = 2).
Example: In maize, incomplete linkage was observed by Hutchinson w.r.t seed coat colour and seed shape. The results show that parental combination of alleles (CS/CS and cs/cs) appear in about 96% cases. The other two are new combinations (Cs/cs and cS/cs) wh ich appear in about 4% cases. Thus, in about 4% cases, crossing over has occurred between linked genes.

Concept Builder
Linkage Group
A set of genes on a chromosome that tend to be inherited (transmitted) which can inherit together from one generation to another generation, constitute a linkage group.
Number of linkage group in an organism is equal to 'n' number of chromosome, e.g., it is 12 in rice, 7 in Pisum, while in human beings, the number of linkage group is 23 in human female and 24 in human male and in Drosophila it is 4.
CROSSING OVER AND RECOMBINATION
Crossing over is a process that produces new combination of genes by interchanging of segments between nonsister chromatids of homologous chromosomes.
The crossing over occurs in between the homologous chromosomes at four stranded or tetrad stage during pachytene of prophase I of Meiosis I.
A condition where an individual heterozygous for two pairs of linked genes (AaBb) possesses the two dominant genes on one member of the chromosome pair and the two recessive on the other, is said to be cis arrangement.

Cis arrangement
If an individual heterozygous for two pairs of linked genes (AaBb) possesses one dominant and one recessive allele of each pair of genes on each member of the homologous pair of chromosome, the arrangement is said to be Trans arrangement.

Trans arrangement
When two genes are located very close to each other in chromosomes, hardly any crossing over can be detected.
The linkage is broken down due to crossing over.
Crossing over will be relatively more frequent if the distance between two genes is more.
Frequency of crossing over can be determined cytologically by counting the number of chiasmata.
The details of the crossing over for two genes A and B and their alleles a and b on the homologous chromosomes are shown in figure.
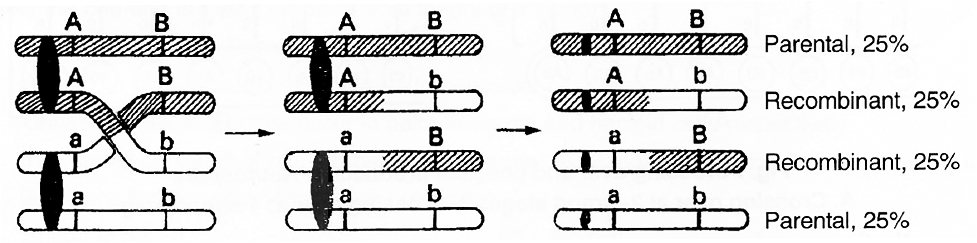
Fig. : Crossing over showing 50 per cent parental and 50 percent recombinants
The crossing over as shown above results in the formation of following four types of cells :

Crossing Over Occurs at Four Stranded Stage
Neurospora (pink mould), an ascomycetous fungus is used to demonstrate that crossing over takes place at 4-stranded stage.
It has following advantages as experimental organism :
(1) It is haploid and there is only one allele at each locus. Hence, dominant-recessive relationship does not interfere with observations and analysis.
(2) The products of single meiosis can be easily analysed.
(3) The products of meiosis occur in the form of 'ordered tetrads', i.e., the eight ascospores formed, are linearly arranged in a sac-like structure called ascus.
If genes A and B are located on same chromosome and undergo independent assortment, the genotype of linearly arranged ascospores can be studied. If crossing over takes place at 2-strand stage, the ascospores would show Ab, Ab, Ab, Ab, aB, aB, aB, aB, (4 Ab + 4 aB) arrangement.
If crossing over takes place at 4-stranded stage, the ascospores would show AB, AB, Ab, Ab, aB aB, ab, ab, (2AB + 2 Ab + 2 aB + 2 ab) arrangement or 2: 4 : 2 arrangement.
Tetrad analysis has demonstrated the presence of such an arrangement and thus, it is now confirmed that crossing over occurs at 4-stranded stage.
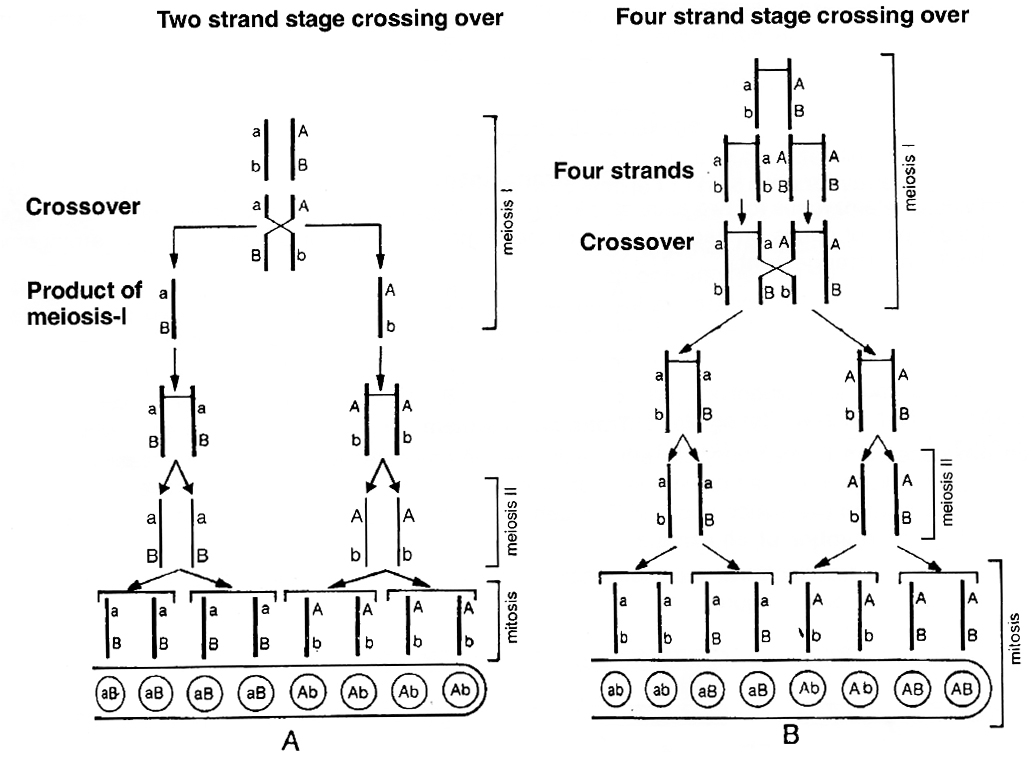
A. Crossing over at 2-strand stage; B. Crossing over at 4-strand stage
Factors affecting crossing over:
(i) Distance between the genes is directly proportional to crossing over.
(ii) Cross over decreases with age.
(iii) X-rays and temperature increases crossing over.
(iv) Centromere and heterochromatin positions decrease crossing over.
(v) One cross over reduces the frequency of other crossover in its vicinity which is called as interference.
Chromosomal Mapping
Crossing over is important in locating the genes on chromosome.
The genes are arranged linearly on the chromosome.
This sequence and the relative distances between various genes is graphically represented in terms of recombination frequencies or cross over values (COV).
This is known as linkage map of chromosome.
Distance or cross over units are called centimorgan (cM) or map unit.
Term centimorgan is used in eukaryotic genetics and map unit in prokaryotic genetics.
Recombination frequency or cross over value = 
The recombination frequency depends upon the distance between the genes.
If the distance between the genes is lesser the chances of crossing over is less and hence recombination frequency is also lesser and vice versa.
So, recombination frequency is directly proportional to the distance between genes.
In any cross, if recombination frequency is 5%.
It means the distance between the genes is 5 map unit.
A.H. Sturtevant suggested that these recombination frequencies can be utilized in predicting the sequence of genes on the chromosome.
On the basis of recombination frequency, he prepared first chromosomal map or genetic map for Drosophila.
Concept Builder
1. T.H. Morgan (1910) developed the theory of the gene as a locus or discrete unit on the chromosomes from his studies on Drosophila melanogaster.
2. Stern (1931) showed that genetic crossing over was accompanied with actual exchange of chromosome segments. He, therefore, gave the cytological proof of crossing over.
3. Map unit is related with frequency of crossing over and not with linkage.
4. C.D. Darlington -Precocity theory of crossing over.
5. J. Belling -Belling's hypothesis and copy choice mechanism of crossing over.
6. Creighton and McClintock -Cytological evidence of crossing over using corn as the material.
7. Pontecarvo (1952) -Defined genes as the ultimate unit of recombination.
8. Janssen -Discovered Chiasmata formation.
sex determination
- Books Name
- A TEXT OF BIOLOGY - CLASS XII
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 12
- Subject
- Biology
SEX DETERMINATION
Sex Chromosomes and Autosomes
Sex chromosomes are those chromosomes which determine the sex of the individual in dioecious or unisexual organisms.
The normal chromosomes, other than the sex chromosomes, of an individual are known as autosomes. Sex chromosomes may be similar in one sex and dissimilar in the other.
The two conditions are respectively called homomorphic (= similar, e.g., XX, ZZ) and heteromorphic (= dissimilar, e.g., XY, ZW).
Individuals having homomorphic sex chromosomes produce only one type of gametes.
They are, therefore, called homogametic (e.g., male birds, human female and Drosophila female).
Individuals having heteromorphic sex chromosomes produce two type of gametes (e.g., X and Y).
They are termed as heterogametic (e.g., female bird, human male and normal Drosophila male).
The factors which control the sex of an organism are under genetic control.
Various mechanism which led to sex determination can be classified into following four categories:
1. Chromosomal mechanism of sex determination.
2. Non -Allosomic genetic sex determination -Fertility factor (plasmid) in bacteria.
3. Genic balance mechanism or X/A balance.
4. Environmental mechanism of sex determination.
1. Chromosomal mechanism of sex determination.
According to this mechanism, there are certain chromosomes known as sex chromosome or heterosome or idiochromosome, which are responsible for sex determination.
This may be of following types:
(i) XX -XY type:
In most insects, plants and mammals including human beings, the females possess two homomorphic (= isomorphic) sex chromosomes, i.e., XX.
The males contain two heteromorphic sex chromosomes, i.e., XY. The Y-chromosome is often shorter and heterochromatic (made of heterochromatin).
Despite differences in morphology, the Y-chromosome pairs with X-chromosome at a certain segment during meiosis.
It, therefore, carries a segment which is homologous with a segment of X-chromosome.
The remaining segment of Y-chromosome is non-homologous and carries only Y-linked or holandric genes, e.g., Testis Determining Factor (TDF).
Human beings have 22 pairs of autosomes and one pair of sex chromosomes.
All the ova (haploid) formed by female are similar in their chromosome type (22 + X). Therefore, females are homogametic.
The male gametes or sperms (haploid) produced by human males are of two types, (22 + X) and (22 + Y).
Human males are therefore, heterogametic.
The two sexes produced in the progeny may have 50 : 50 ratio.
This type of sex-determination was reported in a plant Sphaerocarpus for the first time and is also found in plants like Melandrium and Coccinia.
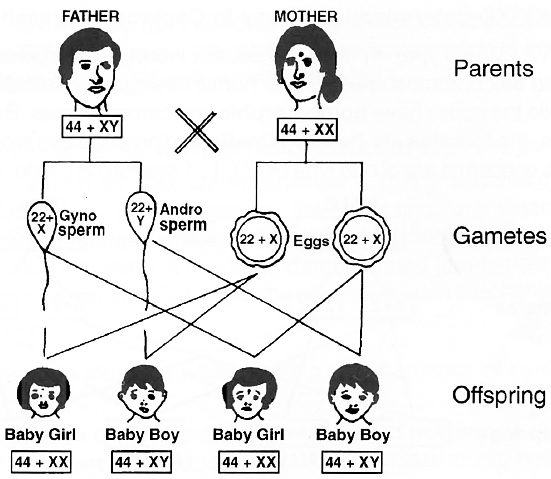
one-half of the sperms carry an X-chromosome and one-half carry a Y-chromosome.
Concept Builder
Barr Body and Y Spots
Barr and Bertram (1949) found a small darkly stained chromatin body (heterochromatic) adhered to the nuclear membrane of nerve cells of female cats but not in male cats.
Later on, Barr (1960) observed consistent presence of these chromatin bodies in epidermal cells of buccal and vaginal mucosa, skin cells etc. in females.
Lyon (1962) suggested that one X chromosome of maternal side becomes coiled, inert and hetero-pycnotic and forms a barr body.
Number of barr bodies is always one less than the total number of X chromosomes in female.
Normal male has no barr body.
In about 5% of polymorphonuclear leucocytes (neutrophils) in women, the heteropycnotic X chromosome occurs as a round pedunculated body attached to the nucleus.
It is called drum stick (Barr body).
A normal female has 1 drum stick per nucleus of neutrophil.
In neutrophils of males drum stick is absent.
Y spots: Males are identified by number of Y spots (Zech 1970). Number of Y spots are equal to number of Y Chromosomes.
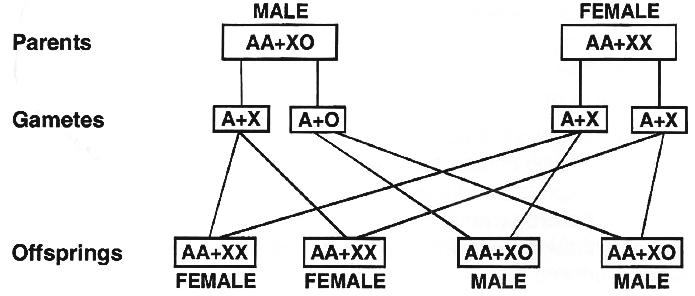
(ii) XX -XO type.
In roundworms, Dioscorea and some insects (true bugs, grasshoppers and cockroaches), the females have two sex chromosomes, XX, while the males have only one sex chromosome, X.
There is no second sex chromosome.
Therefore, the males are designated as XO.
The females are homogametic because they produce only one type of eggs.
The males are heterogametic with half the male gametes carrying X-chromosome while the other half being devoid of it.
The sex ratio produced in the progeny is 1 : 1.
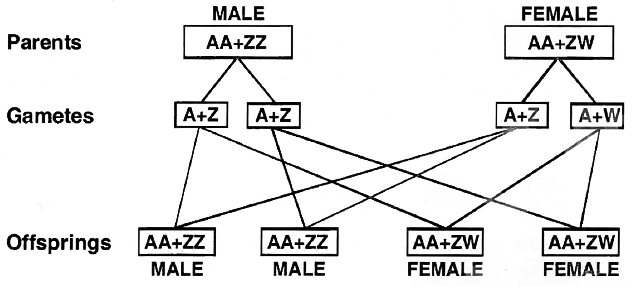
(iii) ZW -ZZ type. (WZ -WW Type).
In birds, fishes, silk worm, Fragaria elatior and some reptiles both the sexes possess two sex chromosomes.
Unlike human beings, the females contain heteromorphic sex chromosomes while the males have homomorphic sex chromosomes.
Because of having heteromorphic sex chromosomes, the females are heterogametic and produce two types of eggs, (A + Z) and (A + W).
The male gametes or sperms are of one type (A+Z). 1 : 1 sex ratio is produced in the offspring.
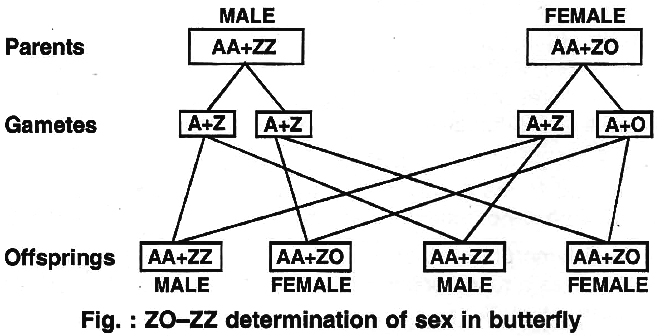
(iv) ZO -ZZ type.
This type of sex determination occurs in butterflies, pigeon, ducks and moths.
It is exactly opposite the condition found in cockroaches and grasshoppers.
Here the females have odd sex chromosome while the males have two homomorphic sex chromosomes.
The females are heterogametic.
They produce two types of eggs, with one sex chromosome (A + Z) and without the sex chromosome (A + 0).
The males are homogametic, forming similar types of sperms (A + Z).
The two sexes are obtained in the progeny in 1 : 1 ratio as both the types of eggs are produced in equal ratio.
Arrhenotoky/Haploid-diploid Mechanism:
This mechanism is found in honey bee. In honey bees, ants and wasps, the egg, if fertilized, gives rise to female fly.
The unfertilized egg develop parthenogenetically into male. So, the female flies are diploid while male flies are haploid.
2. Non-Allosomic genetic sex determination:
Fertility factor of plasmid in bacteria determines sex.
3. Genic Balance or XlA Balance Theory of Sex Determination:
Given by C.B. Bridges. According to him, Y chromosome plays no role in sex determination of Drosophila and it is the ratio between number of X-chromosome and set of autosomes which determines the sex of fly.
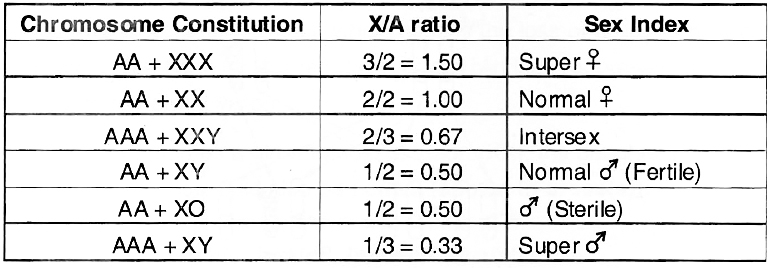
It was concluded that X/A ratio of > 1.0 expresses super femaleness, 1.0 femaleness, below 1.0 and above 0.5 intersexes, 0.5 maleness and < 0.5 supermaleness.
Gynandromorphs : Gynandromorph is a sex mosaic (an individual with one half of the body male and the other half female).
These are common in Silk moth and Drosophila. Gynandromorphism is developed due to accidental loss of X-chromosome from a 2A + XX cell during mitosis.
Gynander : A gynander may be male or female with patches of tissues of other sex on it.
4. Environmental Mechanism of Sex Determination:
This mechanism is observed by F. Baltzer in Bonnelia viridis (marine worm).
In this organism, the sex is undifferentiated in larva.
The larva which settle down in mud, grow up into mature female while those which settle down near the proboscis of female and become parasite develop into male.
It has been demonstrated that female secrete certain hormone which induces sex in larva. Crepidula and Ophryortocha also show such mechanism.
SEX LINKED INHERITANCE
Sex linkage was discovered by Morgan, while working on inheritance of eye colour in Drosophila. He made three types of crosses:
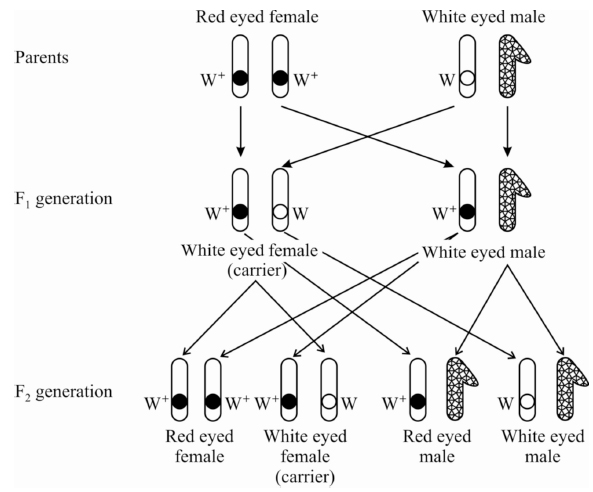
Cross -1 :
The white eyed male (w) was crossed with red eyed (w+) female.
All the flies of F1 generation were found to be red eyed.
F1 flies were allowed to self breed.
In F2 generation, both the traits of red eye and white eye appeared in the ratio 3 : 1 showing that white eye trait is recessive to red eye trait.
Cross -2 :
Red eyed females of F1 generation were crossed with white eyed male.
It is similar to test cross where hybrids are cross bred with recessive parents.
Morgan obtained red and white eyed female as well as male in equal proportions-1 red eyed female: 1 white eyed female: 1 red eyed male: 1 white eyed male.
The test cross indicated that white eye colour was not restricted to the male fly. Red eyed White eyed hybrid female male
traits appear in both males and females in equal proportions
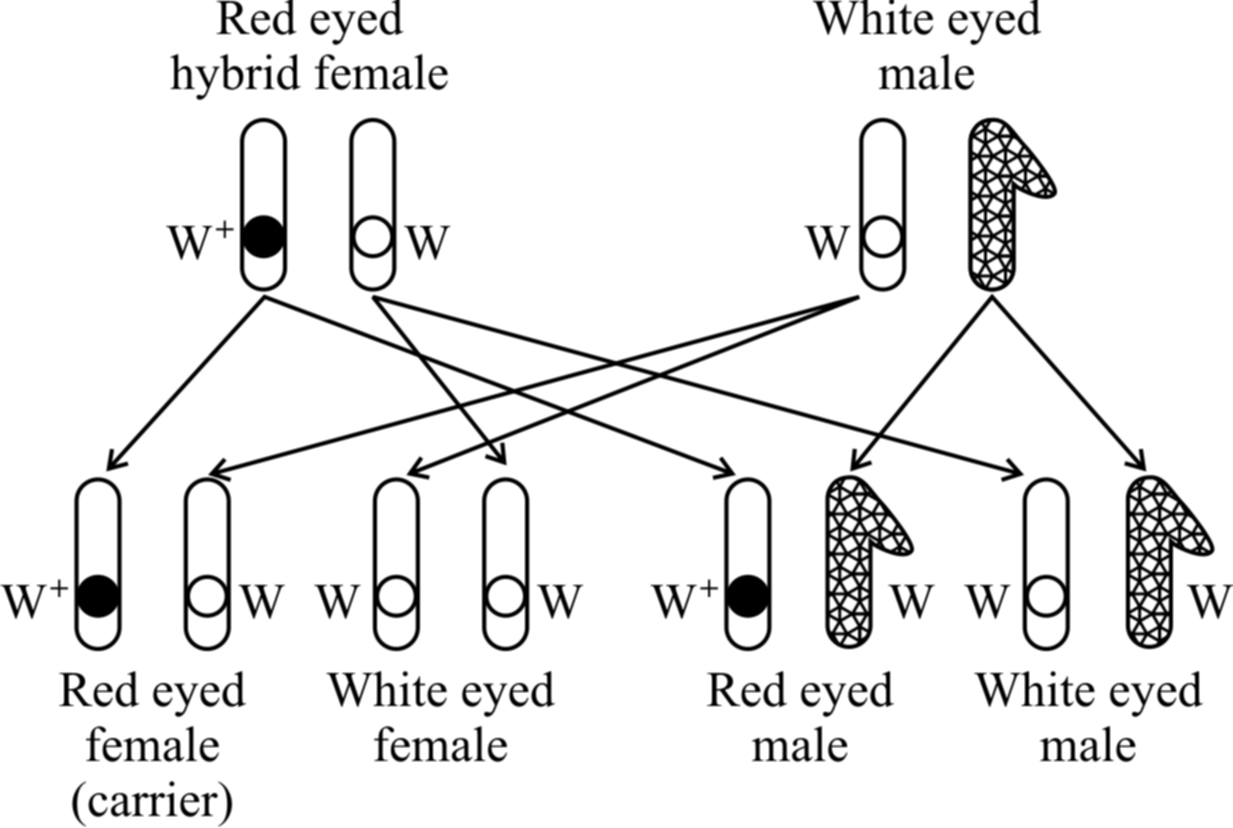
Cross -3 :
White eyed females were crossed with red eyed males. It was a reciprocal of cross 1 and should give the similar result as obtained by Mendel. However, Morgan obtained a surprising result. All the males were white eyed while all the females were red eyed.
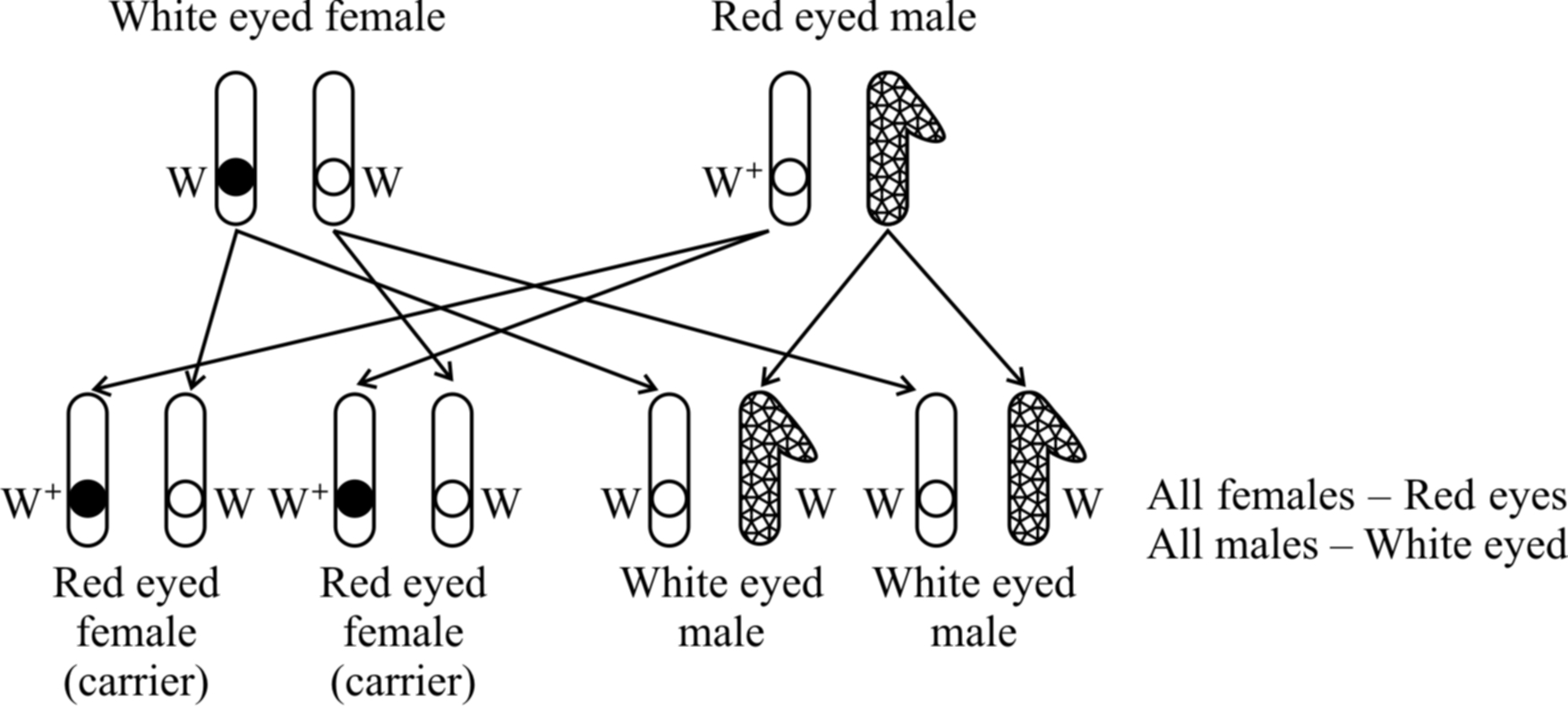
Taking all the crosses into consideration, Morgan came to the conclusion that eye colour gene is linked to sex and is present on the X-chromosome.
X-chromosome does not pass directly from one parent to the offspring of the same sex but follows a criss-cross inheritance, i.e., it is transferred from one sex to the offspring of the opposite sex.
In other words, in criss-cross inheritance a male transmits his traits to his grandson through daughter (Diagynic), while a female transmits the traits to her granddaughter through her son (Diandric).

Sex Linkage in Human Beings
Colour blindness and haemophilia (Bleeder's disease) are two common examples of sex-linked diseases in human beings.
(1) Colour blindness.
This is a human disease which causes the loss of ability to differentiate between red colour and green colour.
The gene for this red-green colour blindness is present on X chromosome. Colour blindness is recessive to normal vision.
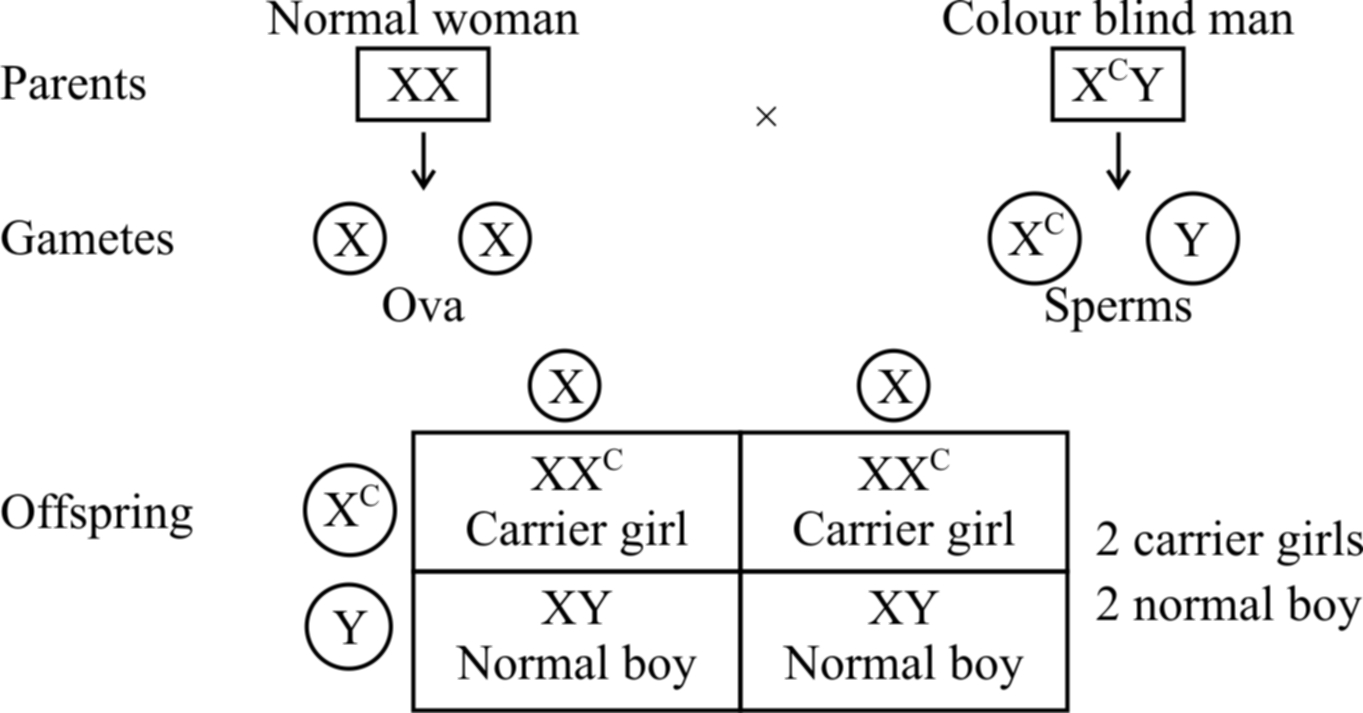
If a colour blind man (XcY) marries a girl with normal vision (XX), the daughters would have normal vision but would be carrier, while sons would also be normal (shown in cross(a)).
Cross (a)
If the carrier girl (heterozygous for colour blindness, XCX) now marries a colour blind man XCY, the offspring would show 50% females and 50% males.
Of the females, 50% would be carrier for colour blindness and the rest 50% would be colour blind.
Of the males, 50% would have normal vision and the 50% would be colour blind (shown in cross (b).
Cross (b)
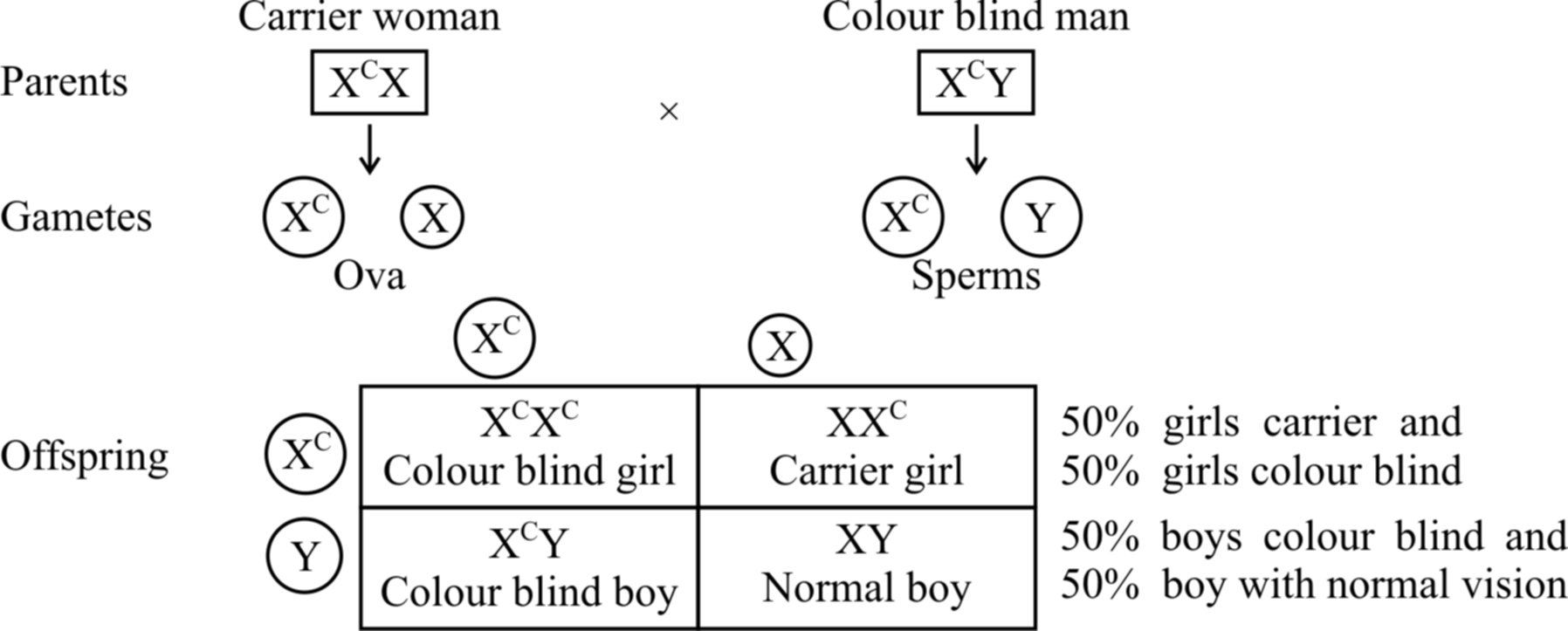
Fig. : Sex-linked inheritance of colour blindness -cross (a) and cross (b)
(2) Haemophilia (Bleeder's disease).
The person suffering from this disease cannot synthesize a normal blood protein called antihaemophilic globulin (AHG) required for normal blood clotting (Haemophilia A - more severe).
Therefore, even a very small cut may lead to continuous bleeding for a long time.
This gene is located on X chromosome and is recessive.
It remains latent in carrier females.
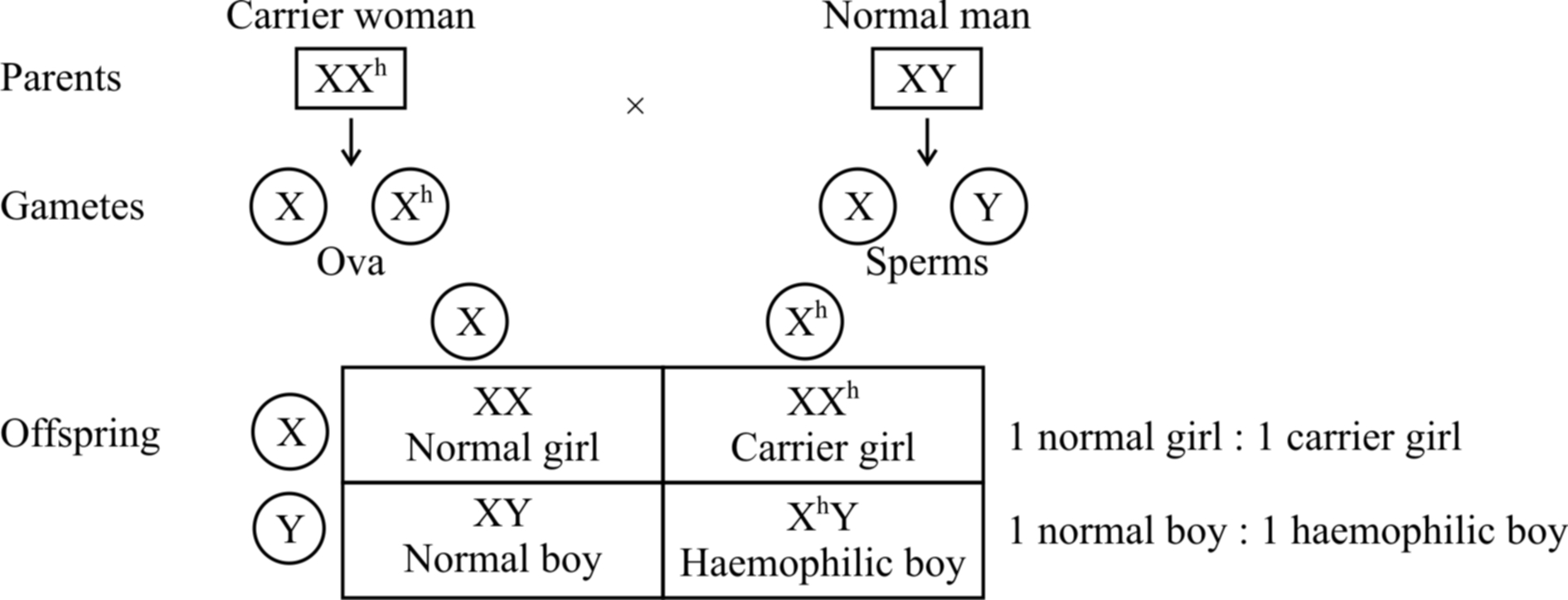
Fig.: Inheritance of haemophilia when the mother is carrier and the father is normal
If a normal man marries a girl who is carrier for haemophilia, the progeny would consist of 50% females and 50% males.
Of the females, 50% would be normal and the rest 50% would be hemophilia carrier.
Of the males, 50% would be normal and the rest would be haemophiliacs.
Haemophilia-B (christmas disease) -plasma thromboplastin is absent, Inheritance is just like Haemophilia A.
Concept Builder
1. C.E. McClung (1902) concluded that some special chromosome (X-chromosome) must be responsible for determination of sex, thus providing first definite evidence for chromosomes theory.
2. Bridges (1916) correlated cytological observations of chromosomes and inheritance of characters.
3. Y-chromosome is male determiner in man but not in Drosophila. However, its accumulation is responsible for motility of sperms in Drosophila.
4. Man is hemizygous for sex chromosomes as he has XY chromosomes.
5. Henking discovered an "X-body" in reproductive cells of firefly.
6. Y-body was discovered by Stevens.
7. Wilson and Stevens -put forward chromosome theory of sex and name X and Y body as X-chromosome and Y-chromosome.
8. Tijo and Levan -Discovered exact number of chromosomes in human beings as 46.
9. F.R. Lillie -Formation of free martins (twins in cattles).
Mutation
- Books Name
- A TEXT OF BIOLOGY - CLASS XII
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 12
- Subject
- Biology
MUTATION
Mutation is sudden, inheritable, discontinuous variation due to change in chromosomes and genes.
Hugo de Vries (1901) one of the discoverers of Mendel's laws, observed two distinct varieties of Oenothera lamarckiana (Evening primrose).
These differed in length of stem, flower form and colour and shape of leaves.
These mutant varieties are now known to have been produced due to chromosomal aberrations.
Seth Wright (1791) is considered to be the first to record point or gene mutation.
He noticed a lamb with unusually short legs.
This short legged breed of sheep was known as ancon breed.
Darwin called this variation as sports.
Bateson (1894) termed them as discontinuous or saltatory variations.
The credit for starting scientific study of mutations goes to Thomas Hunt Morgan (1910).
He is known for his work on fruit fly, Drosophila melanogaster.
He found white eyed mutant of Drosophila and since then about 500 mutations have been observed by geneticists from different parts of the world.
Types of Mutations:
Different classifications of mutations are known, each based on a definite criterion or character.
I. Spontaneous and induced mutations:
These are based upon the agency involved.
Spontaneous mutations. These are natural mutations. They have also been called background mutations. Such mutations occur at a frequency of 1 × 10–5 in nature.
Induced mutations. These have been observed in organisms due to specific factors such as radiations, ultra violet light or variety of chemicals. The agents which induce mutations on their application, are called mutagens or mutagenic agents.
II. On the basis of the type of cells in which mutations occur, there are other two types of mutations.
(a) Somatic mutations. These mutations occur in somatic cells, i.e., body cells or the cells other than germinal cells. The somatic mutations do not have any genetic or evolutionary importance. This is because only the derivatives or the daughter cells formed from the mutated cell will show mutation and not the whole organism.
(b) Germinal mutations. These mutations occur in the gametes or germ cells and are also known as gametic mutations. Such mutations are heritable, and, therefore, are of great evolutionary significance. If the mutations are dominant, these are expressed in the next generation and if recessive, their phenotypic expressions remain suppressed.
III. Forward and backward mutations:
The commonest type of mutation, is the change from normal or wild type to new genotype (recessive or dominant).
Such mutations are called forward mutations.
An organism which has undergone forward mutation, may again develop mutation which restores the original wild-type phenotype.
Such reversions are known as backward mutations or reverse mutations.
Mutations can occur at any stage during the life cycle of an organism.
Types of Mutations:
1. Gene mutation.
It is alteration in the sequences of nucleotide in nucleic acids or any change in the sequence of triplet bases.
If gene mutation arise due to change in single base pair of DNA, it is called point mutation.
Gene mutation occurs by following methods :
(a) Frame-shift mutation (Gibberish mutation)
(i) Deletion : Removal of one or more bases from nucleotide chain.
(ii) Insertion or addition: Addition of one or more bases in a nucleotide chain.
(b) Substitution. The replacement of one base by another.
It is of two types :
(i) Transition: When a purine base (A or G) is substituted by another purine base or pyrimidine base (T or C) is substituted by another pyrimidine base.
(ii) Transversion: Substitution of a purine base with a pyrimidine base or vice versa.
(c) Tautomerization: The purines and pyrimidines in DNA and RNA may exist in several alternate forms or tautomers. Tautomerization occurs through rearrangement of electrons and protons in the molecule. Tautomers show changed base pairing so as to cause change in sequence like AT to CG.
Non-sense Mutation : Such mutations arise when a normal codon, coding for an amino acid is changed into a chain terminating codon (UAG, UAA, UGA) resulting in the production of an incomplete polypeptide.
Nonsense mutations rarely go unnoticed because the incomplete or shorter protein formed, is generally inactive.
Mis-sense mutation: Change in base in a codon, producing a different amino acid at the specific site in a polypeptide. In mis-sense mutation, the change in one amino acid is frequently compatible with some biological activity, e.g., sickle cell anaemia.
2. Chromosomal mutation (Chromosomal Aberrations) :
The change in the chromosome morphology is called chromosomal aberration. Structural changes in the chromosomes take place during meiosis. There are four types of chromosomal rearrangements:
(a) Deficiency :
Loss of a terminal segment of chromosome or Deletion -It is due to loss of a intercalary part of chromosome, e.g., Cri-du-Chat syndrome (short arm of chromosome 5 loses a part).
(b) Duplication:
Occurs due to addition of a part of chromosome so that a gene or set of genes is represented twice, e.g., Barr eye in Drosophila.
(c) Translocation :
It involves shifting of a part of one chromosome to another non-homologous chromosome. So new recombinant chromosomes are formed, as this induces faulty pairing of chromosomes during meiosis. An important class of translocation having evolutionary significance is known as reciprocal translocation or segmental interchanges, which involves mutual exchange of chromosome segments between non-homologous chromosome, i.e., illegitimate crossing over. Chronic myelogenous leukemia (CML) occurs due to translocation of segment of long arm from chromosome 22 to chromosome 9. Chromosome 22 is called Philadelphia chromosome.
(d) Inversion:
Change in linear order of genes by rotation of a section of chromosome by 180°. Inversion occurs frequently in Drosophila as a result of X-ray irradiation. They may be of two types:
(i) Paracentric : Inversion without involving centromere (Inverted segment does not carry centromere).
(ii) Pericentric: Inversion involving centromere.
3. Genomatic mutation or numerical changes in chromosome number :
It is of two types:
(A) Aneuploidy:
In aneuploidy, any change in number of chromosomes in an organism would be different than the multiple of basic set of chromosomes. It results due to failure of segregation of chromatids during cell division cycle. There would be following two possibilities:
(a) Hypoploidy:
This arises due to loss of one or more chromosomes or pairs of chromosome. Thus, the following conditions are likely to be produced:
(i) Monosomy (2n -1) : It is the result of loss of one chromosome for a homologous pair. Its other variant is double monosomy, i.e., (2n -1 -1).
(ii) Nullisomy (2n -2) : It is the result of loss of a complete homologous pair of chromosome.
(b) Hyperploidy: This arises due to addition of one or more chromosomes or pairs of chromosome. The following conditions are thus likely to be produced :
(i) Trisomy (2n + 1) : In this type, a single chromosome is added to the chromosome set. The trisomics were obtained for the first time in Datura stramonium (Jimson weed) by A.F. Blakeslee and his co-workers (1924). In human beings, Mongolism or Down's syndrome is due to trisomy of chromosome 21 (2n + 1 or 46 + 1, i.e., chromosome 21 is present 3 times). Others are Patau syndrome (due to trisomy of 13th) and Edward's syndrome (due to trisomy of 18th chromosome).
(ii) Tetrasomy (2n + 2) : It is the result of addition of a complete homologous pair of chromosomes (i.e., 2-chromosomes).
(B) Euploidy:
In euploidy, any change in the number of chromosomes is the multiple of the number of chromosomes in a basic set or it occurs due to variation in one or more haploid sets of chromosomes. Accordingly, these may be haploid and polyploid.
(a) Haploidy : One set of chromosomes. Haploids are better for mutation experimental studies, because all mutations either dominant or recessive can express immediately in them, as there is only one allele of each gene present in each cell.
(b) Polyploidy: Failure of cytokines is after telophase stage of cell division results in an increase in a whole set of chromosomes in an organism and this phenomenon is known as polyploidy. Polyploids are of particular importance and are, therefore, discussed below. Polyploids fall into two major categories:
(i) Autopolyploids have the same basic set of chromosomes multiplied more than twice, e.g., AAA -autotriploid, AAAA -Autotetraploid. Autopolyploids with odd number of chromosomes are seedless but show Gigantism (large size) or gigas effect (more yield and more adaptability), but are odd numbered so can only be propagated vegetatively, e.g., Banana, Pineapple. Naturally occurring autotriploids are known in banana, grapes, sugar beet, tomato and watermelons. Similarly, autotetraploids occur amongst apples, berseem, corn etc.
Autopolyploids can also be produced artificially by treating the seeds, seedlings or axillary buds with an alkaloid called colchicine. It is extracted from corm of Colchicum autumnale. The treatment produces doubling of chromosomes.
(ii) Allopolyploids are hybrids whose chromosome sets are derived from two different genomes.
Let A and B be considered as different genomes. The two diploid organisms should have AA and BB chromosome sets. The autotetraploid can now be represented as AAAA while allotetraploid is represented as AABB.
Such polyploids are the result of doubling of chromosomes in F1 hybrids derived from two different or related species.
The commonest example of allopolyploidy is Raphanobrassica developed by Russian geneticist G.O. Karpechenko (1927). A cross was made between Brassica oleracea (2n = 18) and Raphanus sativus (2n = 18). F1 hybrid produced was sterile, because chromosome sets of both these plants were dissimilar and could not pair during meiosis. However, some fertile plants with 2n = 18A + 18B (18 bivalents) were found amongst these.
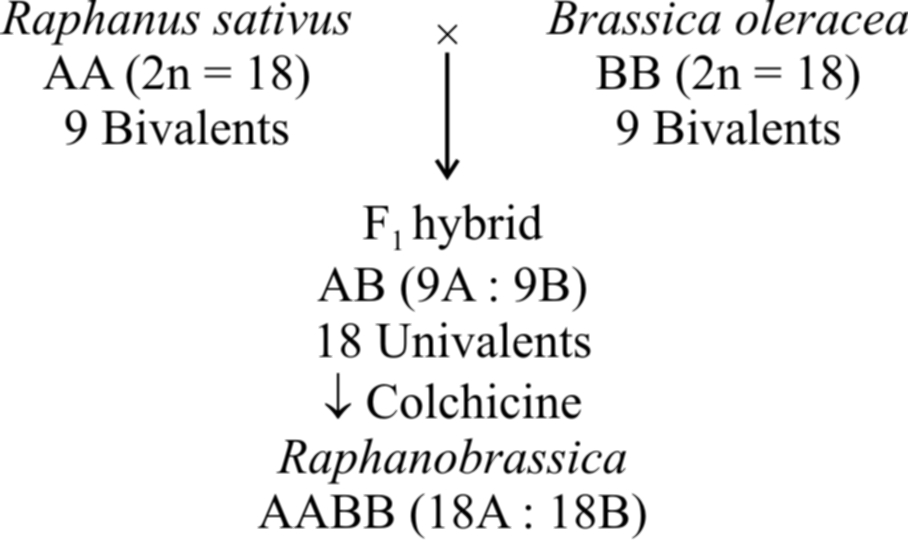
Another example is a man made cereal -Triticale, produced by (i) crossing Triticum durum (2n = 28) with Secale cereale (2n = 14) and then treating F1 hybrid with colchicine to obtain hexaploid Triticale and (ii) crossing Triticum aestivum (2n = 42) with Secale cereale (2n = 14) and then treating F 1 hybrid with colchicine to produce octaploid Triticale.
Mutagens
Mutations can be artificially produced by certain agents called mutagens or mutagenic agents. Following are two major types of mutagens :
(1) Physical mutagens
(2) Chemical mutagens
(1) Physical mutagens:
Radiations are the most important physical mutagens.
H.J. Muller who used X-rays, for the first time, to increase the rate of mutation in Drosophila, opened an entirely new field in inducing mutations.
So, Muller is considered as father of Actinobiology.
The main source of spontaneous mutations are the natural radiations coming from cosmic rays of the sun.
The spectrum of wavelengths that are shorter (i.e., of higher energy) than the visible light can be subdivided into following two groups.
(a) Ionizing radiations
(b) Non-ionizing radiations
They occur in small amounts in the environment and are known as background radiations.
The following are biological effects of radiations :
(a) Effects of ionizing radiations: The ionizing radiations include X-rays, -rays, -rays and -rays. Ionizing radiations cause breaks in the chromosome. These cells then show abnormal cell divisions. If these include gametes, they may be abnormal and even die prematurely. Different types of cancers may result due to radiations. The frequency of induced mutations is directly proportional to the doses of radiations.
(b) Effects of non-ionising radiations : The non-ionizing radiations have longer wavelengths but carry lower energy. This energy is insufficient to induce ionization. Therefore, non-ionizing radiations such as UV light do not penetrate beyond the human skin . Thymine (pyrimidine) dimer formation is a major mutagenic effect of UV rays that disturbs DNA double helix and thus, DNA replication.
(2) Chemical mutagens:
Large number of chemical mutagens are now known. These are more injurious than radiations. The first chemical mutagen used was mustard gas by C. Auerbach et. al. during world war II.
Chemical mutagens are placed into two groups:
(a) Those which are mutagenic to both replicating and non-replicating DNA such as nitrous acid,
(b) Those which are mutagenic only to replicating DNA, such as acridine dyes and base analogues.
Following are the effects of some of the chemical mutagens:
(i) Nitrous acid: It is mutagenic to both replicating and non-replicating DNA. It acts directly by oxidative deamination on A, G and C bases which contain amino groups. A is deaminated to hypoxanthine which is complementary to cytosine. G is converted to xanthine which pairs with C. Cytosine is converted to U which pairs with A
(ii) Acridines: Acridines and proflavins are very powerful mutagens. These can intercalate between DNA bases and interfere with DNA replication, producing insertion or deletion or both of single bases respectively. This induces frame shift-mutations or gibberish mutation, e.g., Thalassaemia.
(iii) Base analogues: These have structures similar to the normal bases and are incorporated into DNA only during DNA replication. Base analogues cause mispairing and eventually give rise to mutations. The base analogues may either be natural or artificial. Natural base analogues include 5-methyl cytosine, 5-hydroxymethyl cytosine, 6-methyl purine etc.
The most commonly used artificial base analogues are 5-bromouracil and 2-aminopurine. 5-bromouracil is a structural analogue of thymine. It gets incorporated into the newly replicated DNA in place of thymine (T). 2-aminopurine is an artificial base analogue of adenine. It acts as a substitute of adenine (A) and can also pair with cytosine (C).
CYTOPLASMIC INHERITANCE
Some self replicating genes (DNA) are present in the cytoplasm (mitochondrial DNA and chloroplast DNA) also.
These are called plasmagenes and all the plasmagenes together constitute plasmon (like genome).
The inheritance of characters by plasmagenes is called extranuclear or extrachromosomal inheritance.
Certain most important examples of extranuclear inheritance in eukaryotes are following :
Maternal inheritance:
The amount of nuclear hereditary material contributed by the two sexes is almost equal but the cytoplasm in egg is always much more than that of the sperm.
So, in extranuclear inheritance, contribution of female parent is more.
This is called maternal inheritance.
The evidence of maternal inheritance is the coiling of shell in snails.
Concept Builder
Shell Coiling in Snails
Shell coiling in Limnaea peregra, a fresh water snail, is of two types, dextral ( =clockwise) and sinistral (anticlockwise).
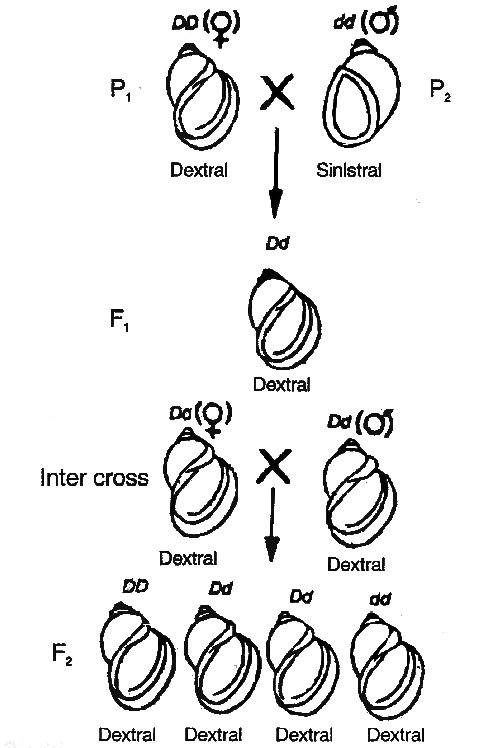
 dextral (DD) and
dextral (DD) and  sinistral (dd) snails
sinistral (dd) snailsDextral coiling is controlled by a dominant nuclear gene called D while sinistral coiling is due to a recessive gene d.
Since, zygote receives whole of its cytoplasm from the egg, the direction of shell coiling in the offspring is governed by cytoplasm of the mother.
When dextral female (DD) is crossed with sinistral male (dd), all the offsprings of F1 generation (Dd) have dextral coiling.
If sinistral female (dd) is crossed with dextral male (DD), the offspring have Dd genotype but coiling is sinistral.
When the latter are self-bred, three types of genotypes are produced1DD, 2Dd and 1dd.
However, shell coiling in all is dextral because the mother has Dd genotype.
Organelle inheritance:
The DNA is present in mitochondria and chloroplast which controls the inheritance of some characters.
A well known example of the characters controlled by chloroplasts is plastid inheritance in Mirabilis jalapa (4 O'clock plant), discovered by Correns.
Other examples of organellar inheritance are iojap inheritance in maize, inheritance of poky (imbalance in the mitochondrial physiology) in the fungus Neurospora crassa and Petite in yeast, a mitochondrial character.
Cytoplasmic male sterility in maize, is also a function of defective mitochondria.
Genetic disorders
- Books Name
- A TEXT OF BIOLOGY - CLASS XII
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 12
- Subject
- Biology
GENETIC DISORDERS
Pedigree Analysis :
A record of inheritance of certain genetic traits for two or more generations presented in the form of a diagram or family tree is called pedigree.
Parents are shown by horizontal line while their offsprings are connected to it by a vertical line.
The offsprings are also shown in the form of a horizontal line below the parents and numbered with arabic numerals.
Pedigree analysis is study of pedigree for the transmission of particular trait and finding the possibility of absence or presence of that trait in homozygous or heterozygous state in a particular individual.
It is useful for the genetic counsellors to advise intending couples about the possibility of having children with genetic defects like haemophilia, colour blindness, alkaptonuria, phenylketonuria, thalassaemia, sickle cell anaemia (recessive traits), brachydactyly, myotonic dystrophy and polydactyly (dominant traits).
Pedigree analysis indicates that Mendel's principles are also applicable to human genetics with some modifications found out later, like quantitative inheritance, sex linked characters and other linkages.
Symbols used in Pedigree analysis :
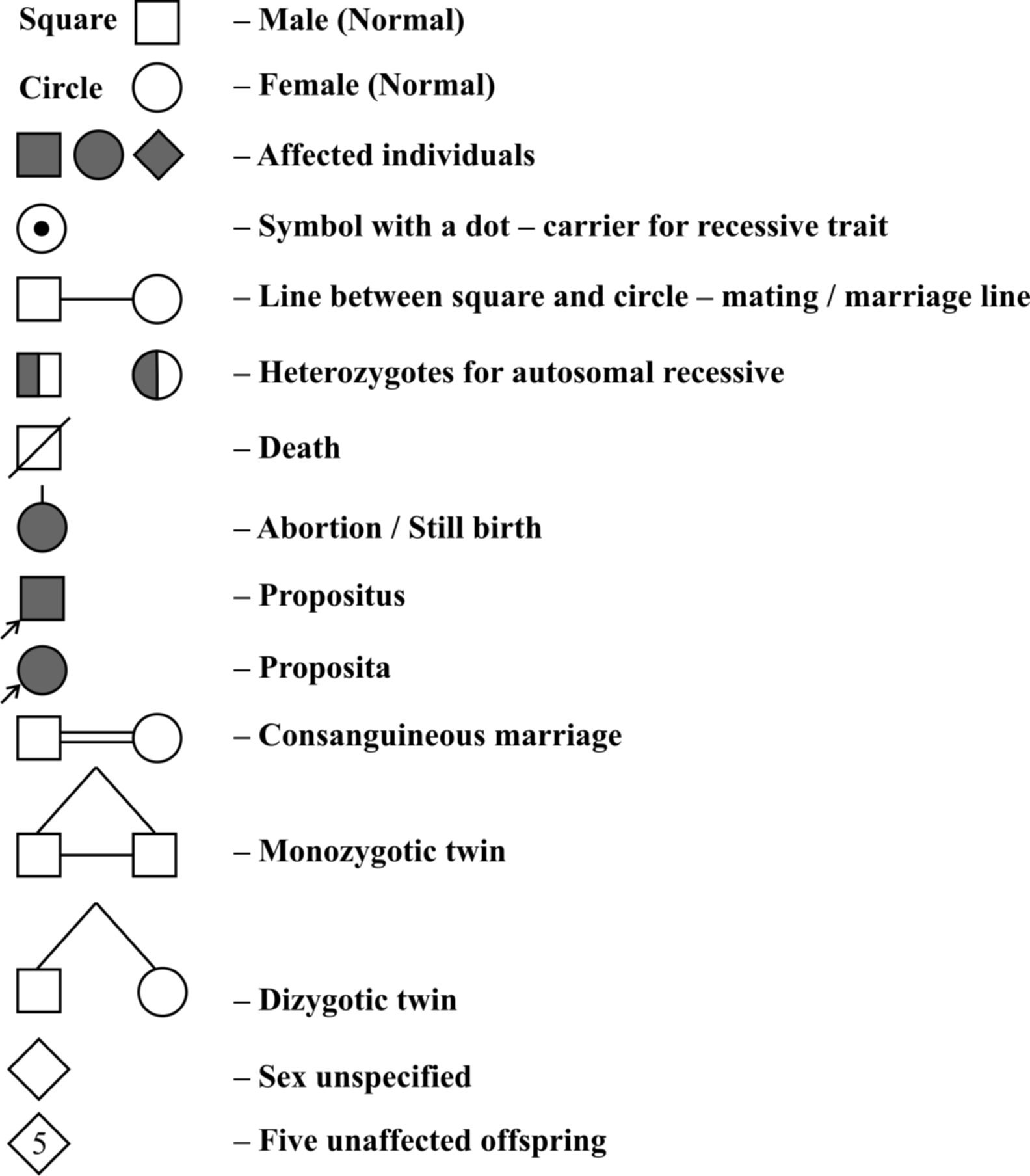
Proband is person from which case history starts. If it is male, it is called propositus, if it is female it is called proposita.
Example 8 : In the pedigree given below, indicate whether the shaded symbols indicate dominant or recessive allele. Also give genotype of the whole pedigree.
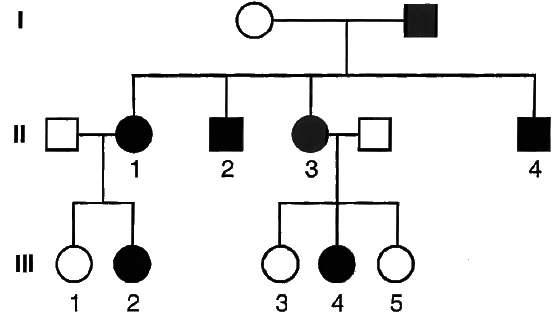
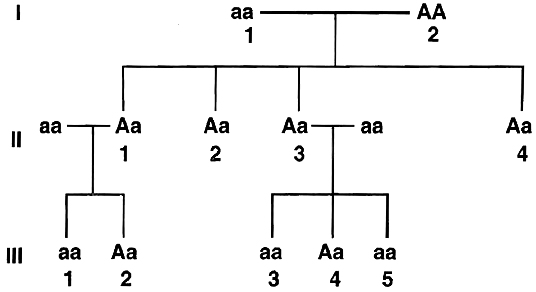
Solution: Since the shaded symbol appears in all the offsprings, father must be homozygous dominant while the mother homozygous recessive (AA × aa = all Aa) because in all other cases this possibility is absent (Aa × aa = 2Aa + 2aa; aa × AA = all Aa; aa × Aa = 2aA + 2aa). All the members of II generation will, therefore, be heterozygous (Aa). This is further confirmed by marriage of II-1 with homozygous recessive (Aa × aa = 2Aa, 2aa) and bearing children of both the parental types. Marriage of II-3 with the homozygous recessive can produce both recessive and heterozygotes as are present here.
MENDELIAN DISORDERS
Sickle-Cell Anaemia
It is an autosomal recessive disorder. In this disorder, the RBCs become sickle shaped under low O2 concentration.
The affected persons die young.
Other heterozygous for this trait are having normal phenotype and long lived.
The disease is due to base substitution of sixth codon in the gene coding for chain of haemoglobin.
The middle base of a DNA triplet coding for the amino acid glutamic acid is mutated so that the triplet now codes for valine instead.
The mutant haemoglobin molecule undergoes polymerisation under low O2 tension causing the change in the shape of RBC from biconcave disc to elongated sickle like structure.
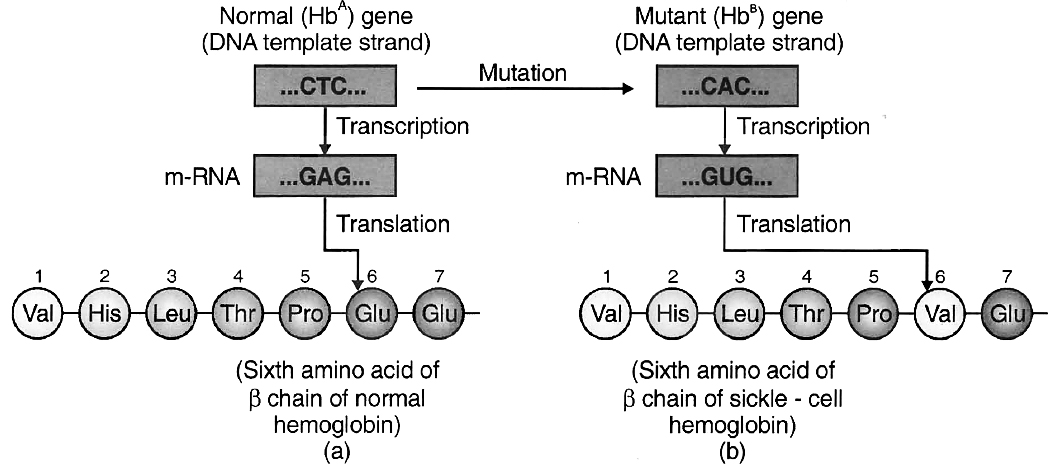
(a) from a normal individual; (b) from an individual with sickle-cell anaemia.
Thalassaemia
It is recessive autosomal disease caused due to reduced synthesis of or polypeptide of haemoglobin. thalassaemia is a major problem, individuals suffering from major thalassaemia often die before ten years of age.
Phenylketonuria
Recessive autosomal disorder (Chromosome 12) related to phenylalanine metabolism to tyrosine. This disorder is due to absence of a liver enzyme called phenylalanine hydroxylase. Due to lack of this enzyme, phenylalanine follows another pathway and gets converted into phenylpyruvic acid. This phenyl pyruvic acid upon accumulation in joints causes arthritis and if it hits the brain, then it causes mental retardation known as phenyl pyruvic idiocy. These are also excreted through urine because of poor absorption by kidney.
Cystic Fibrosis
It is an autosomal recessive disorder common among Caucasian Northern Europeans. Persons suffering from this disease are having extremely salty sweat. It is due to decreased Na+ and Cl– reabsorption in the ducts. Disease is due to a gene present on chromosome 7. Due to a defective glycoprotein, thick mucus develops in pancreas and lungs and formation of fibrous cyst occurs in pancreas.
Huntington's Chorea
It is an autosomal dominant disorder. The gene responsible for this disorder is present on chromosome 4. Disease is characterised by gradual degradation of brain tissue in the middle age and consequent shrinkage of brain.
Alzheimer's Disease
This autosomal recessive disease results in mental deterioration (loss of memory, confusion, anxiety) and ultimately the loss of functional capacities. The disease is due to deposits of -amyloid, a short protein in brain which results in degradation of neurons. It involves two defective alleles located on chromosome number 19 and 21. This disease is common in Down's syndrome.
Myotonic dystrophy is due to a dominant autosomal mutant gene located on the long arm of chromosome 19. Mild myotonia -atrophy and weakness of the musculature of the face and extremities, is most common.
Other Mendelian disorders :
(i) Alkaptonuria (Garrod, 1908) - Due to deficiency of oxidase enzyme.
(ii) Albinism (Chromosome 11) - Absence of tyrosinase
(iii) Tay-Sach's disease (Chromosome 15) - Absence of hexosaminidase B.
(iv) Gaucher's disease (Chromosomes 1) - Due to the inhibition of glucocerebrosidase enzyme action which leads to accumulation of cerebroside.
Other abnormalities due to autosomal dominant gene mutation
(i) Polydactyly - Presence of extra fingers and toes
(ii) Brachydactyly - Abnormal short fingers and toes
Abnormalities due to sex linked (X-linked) recessive gene mutation
(i) Haemophilia A - Due to lack of antihaemophilic-globulin.
Haemophilia B - Due to lack of plasma thromboplastin
(ii) Red-green colour blindness - Daltonism
Protanopia - Red colour blindness
Tritanopia - Blue colour blindness
Deuteranopia - Green colour blindness
(iii) Muscular dystrophy - Due to non-synthesis of protein dystrophin
Deterioration of muscles at an early stage
(iv) Lesch Nyhan syndrome -Deterioration of nervous system
Due to HGPRT deficiency (Hypoxanthin guanine
phosphoribosyl transferase)
CHROMOSOMAL DISORDERS
A. Autosomal abnormalities (Due to mutation in body chromosome)
(i) Down's Syndrome -It occurs due to trisomy of 21st chromosome. The affected individual is short statured with small round head, furrowed tongue and partially open mouth. Palm is broad with characteristic palm crease and mental retardation. Physical and psychomotor development is retarded.
(ii) Edward's syndrome - Trisomy of 18th chromosome
(iii) Patau's syndrome - Trisomy of 13th chromosome
(iv) Cri du chat syndrome - Due to deletion in short arm of 5th chromosome.
B. Allosomal or Sex Chromosomal Disorder
(i) Klinefelter's Syndrome - It occurs due to the trisomy of X-chromosome in male, resulting into a karyotype of 47, (44 + XXY). Individuals have long legs, sparse body hair, small prostrate gland, small testes, reduced mental intelligence and enlarged breasts (Gynaecomastia). Such individuals are sterile.
(ii) Turner's Syndrome - It is caused due to absence of one of the X-chromosome in female i.e. 45 with chromosome complement 44 + XO. Such females are sterile with undeveloped breast, short stature, reduced ovaries & absence of menstrual cycle.
(iii) Super female -AA + XXX, AA + XXXX
(iv) Jacob's syndrome or Super male -AA + XYY, also called as criminal syndrome.
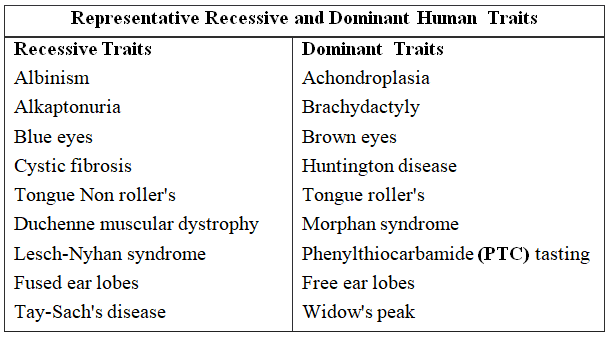
Population Genetics
Hardy Weinberg equation is applied to know the distribution of traits and frequency of autosomal dominant recessive gene distribution in the entire population.
p = Dominant gene/allele
q = Recessive gene/allele
p + q = 1
(p + q)2 = p2 + q2 + 2pq = 1
 (p2 + 2pq) = Frequency of dominant trait.
(p2 + 2pq) = Frequency of dominant trait.
 (q2) = Frequency of recessive trait.
(q2) = Frequency of recessive trait.
Concept Builder
1. Archibald Garrod discovered "Inborn errors of metabolism in humans" -phenylketonuria, alkaptonuria and albinism.
2. Muscular dystrophy and haemophilia causing genes are sex-linked recessive lethal genes.
3. The symptomatic treatment of genetic diseases of man is called Euphenics.
4. Improvement of human race by improving environmental conditions is called Euthenics.
5. Improvement of the future qualities of mankind by selective breeding is called Eugenics.
6. F. Galton -Term "Eugenics".
7. Lejeune -Discovered Cri-du-chat syndrome
8. Darligton (1939) -The genes present in the cytoplasm are called plasmagenes.
