 ACME SMART PUBLICATION
ACME SMART PUBLICATION
The Root
Chapter 5
Morphology of flowering plants
Angiosperms have a wide range of exterior forms or morphology, but they all have roots, stems, leaves, flowers, and fruits in common. Flowers and fruits may be present. The root system of a flowering plant is underground, whereas the component above ground is called the shoot system.
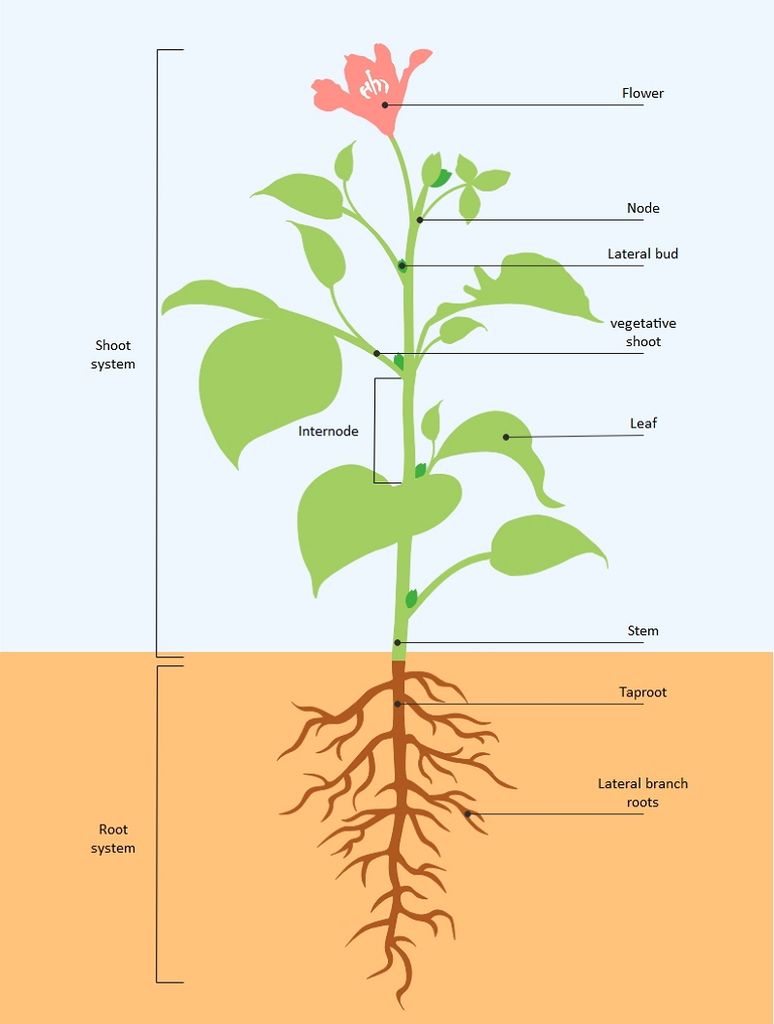
The Root
All vascular plants have roots, which are a crucial subterranean component. This portion of the plant is primarily responsible for attaching it to the ground and absorbing the soil's necessary mineral components, nutrients, and water. It can also be used to keep food.
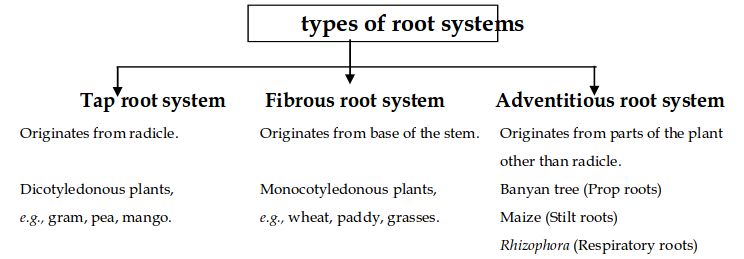
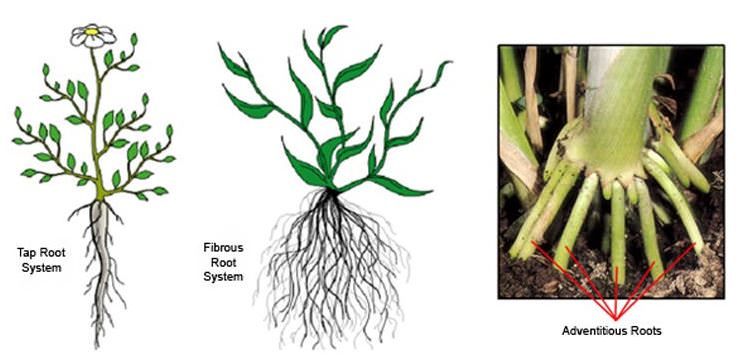
Types of roots:In the majority of dicotyledonous plants, direct radicle elongation results in the creation of the main root that grows inside the soil. It has lateral roots of several orders, referred to as secondary, tertiary, and so on. This is called the taproot system, as shown in the mustard plant. It is made up of the major roots and their branches. The primary root of monocotyledonous plants is short-lived and is replaced by a vast number of roots.As shown in the wheat plant, these roots grow from the base of the stem and form the fibrous root system.
Adventitious roots emerge from portions of the plant other than the radicle in some plants, such as grass, Monstera, and the banyan tree. Absorption of water and nutrients from the soil, appropriate anchorage of plant parts, storage of reserve food material, and synthesis of plant growth regulators are the key tasks of the root system.
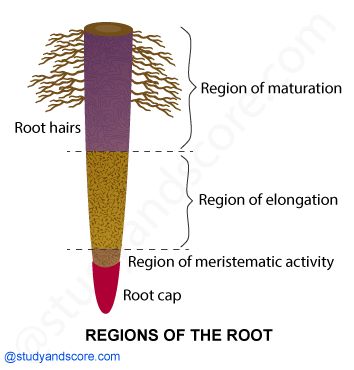
Regions of a root:The root cap is a thimble-like structure that covers the root at its tip. It shields the root's fragile apex as it travels through the soil. Meristematic activity occurs a few centimeters above the root cap. This region's cells are tiny, thin-walled, and have thick protoplasm. They repeatedly divide. The cells closest to this region experience rapid elongation and expansion, and they are responsible for the root's length growth. The elongation region is the name given to this area. The elongation zone's cells mature and differentiate throughout time. As a result, the zone proximal to the region of elongation is known as the maturation zone.Root hairs are incredibly tiny and fragile thread-like structures formed by epidermal cells in this region. The soil's water and minerals are absorbed by these root hairs.
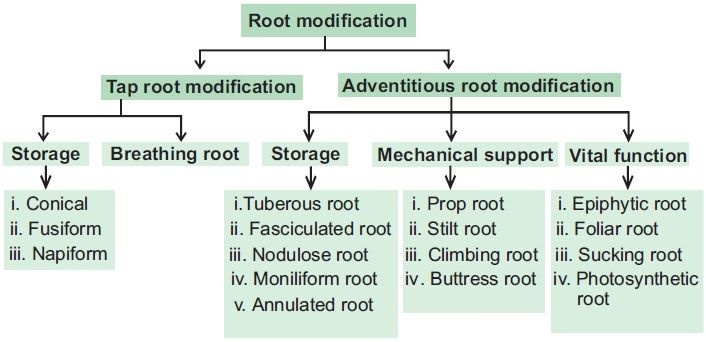
Modifications of a root:Some plants' roots change shape and structure, allowing them to perform roles beyond water and mineral absorption and conduction. They've been altered to provide support, food storage, and breathing. Carrot, turnip, and sweet potato adventitious roots are used for the storage of food in plants. Prop roots are the hanging structures that support a banyan tree. Similarly, maize and sugarcane stems have supporting roots that emerge from the lower nodes of the stem. They are called Stilt roots. Many roots emerge from the ground and climb vertically upwards in plants like Rhizophora that flourish in swampy environments. Pneumatophores are roots that aid in the acquisition of oxygen for breathing.

The Stern
The Stem
A stem is one of a vascular plant's main structural axes. It provides support for leaves, flowers, and fruits, distributes the water and dissolved chemicals between the roots and the shoots in the xylem and phloem, stores nutrients, and generates new living tissue. The stem is the section of the axis that ascends and bears branches, leaves, flowers, and fruits. It grows from the plumule of a sprouting seed's embryo. Nodes and internodes are found on the stem. Nodes are the parts of the stem where leaves are born, whereas internodes are the parts between two nodes. The stem produces buds that are either terminal or axillary. When young, the stem is usually green, but as it matures, it becomes woody and dark brown.
The stem's primary job is to spread out branches containing leaves, flowers, and fruits. Water, minerals, and photosynthates are all carried by it. Some stems have multiple functions, including food storage, support, protection, and vegetative propagation.
Modifications of a Stem: Stems havebeen altered to do various tasks. In potato, ginger, turmeric, zaminkand, and Colocasia, underground stems have been adapted to store food. They also serve as perennation organs, allowing them to survive in environments that are not conducive to growth. Stem tendrils, which emerge from axillary buds and are slender and spirally coiled, assist plants in climbing, such as cucumbers, pumpkins, and melons. Stem axillary buds can also be transformed into woody, straight, and pointed thorns.
Many plants, such as Citrus and Bougainvillea, have thorns. They keep animals from eating the plants. Arid-climate plants change their stems into flattened (Opuntia) or fleshy cylindrical (Euphorbia) forms.They have chlorophyll and are capable of photosynthesis.
Some plants, such as grass and strawberry, stretch underground stems to new niches, and new plants emerge as older sections die. A slender lateral branch emerges from the base of the main axis in plants like mint and jasmine, and after growing aerially for a time, arch downwards to touch the ground.
Aquatic plants like Pistia and Eichhornia have a lateral branch with short internodes and each node containing a rosette of leaves and a tuft of roots. The lateral branches of bananas, pineapples, and Chrysanthemums begin from the main stem's basal and underground portion, develop horizontally beneath the soil, and then emerge obliquely upward, giving rise to leafy shoots.
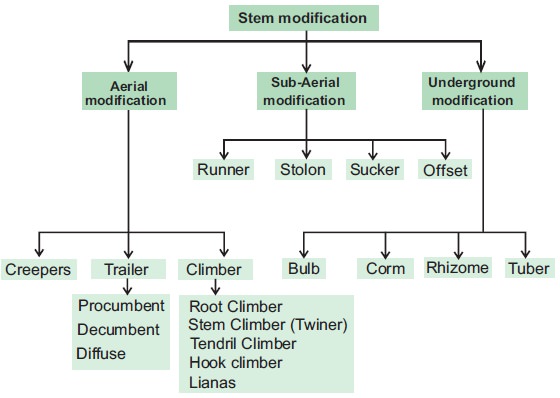
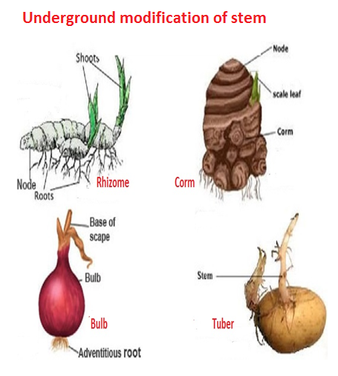

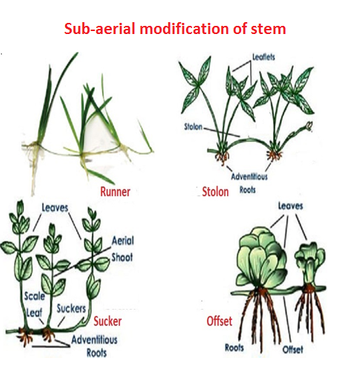
Figure 6(b): Stem modifications
The Leaf
The Leaf
The organ of plant that generates the major lateral appendage on the stem of vascular plants is referred to as a leaf. The leaf is a lateral, flattened structure that grows from the stem. It grows at the node and produces a bud in the axil. A branch arises from the axillary bud. Shoot apical meristems give rise to leaves, which are organized in an acropetal order. They are the most essential photosynthetic vegetative organs. It's also known as the plant's kitchen or food factory. This is due to the fact that they are the primary organ responsible for photosynthesis, which is how the plant obtains its energy or food. The presence of chlorophyll gives them their green color.
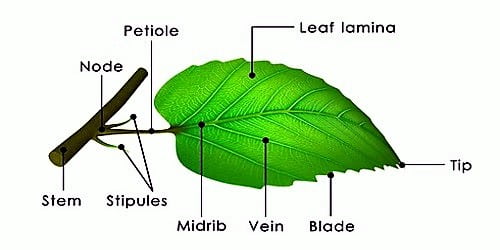
The leaf base, petiole, and lamina are the three main sections of a normal leaf. The leaf base connects the leaf to the stem, while stipules are two lateral tiny leaf-like structures. In monocotyledons, the leaf base develops into a sheath that partially or completely covers the stem. The pulvinus occurs when the leaf base of some leguminous plants becomes inflated. The petiole aids in keeping the blade lit. Leaf-blades flap in the breeze, cooling the leaf and delivering fresh air to the surface, thanks to long thin flexible petioles. The leaf blade, or lamina, is the extended green section of the leaf with veins and veinlets. There is frequently a conspicuous middle vein, referred to as the midrib.Veins give the leaf blade structure and serve as routes for water, minerals, and food components. The shape, edge, apex, surface, and extent of lamina incision vary amongst leaves.
Venation: Venation is the arrangement of veins and veinlets in the lamina of a leaf. The venation is called reticulate when the veinlets create a network. Examples: Hibiscus, papaya, leaves of Tulsi, Coriander, China Rose, and Mangifera. A type of reticulate venation pattern in which the secondary veins run parallel to each other from the midrib toward the margin is called pinnate venation. Examples include Alder, Chestnut, and Oak. Palmate is a venation pattern in which several main veins radiate outward from the base of the leaf. Examples are, Maple, Tulip etc. The venation is called parallel when the veins inside a lamina run parallel to one other. Example: banana, grass, and wheat. Most dicotyledonous plants have reticulate venation on their leaves, but most monocotyledons have parallel venation.A vascular arrangement in leaves such that the veins are forked, with each vein dividing at intervals into smaller veins of approximately equal size is called Dichotomous venation, for example, Ginkgo biloba.

Types of leaves:
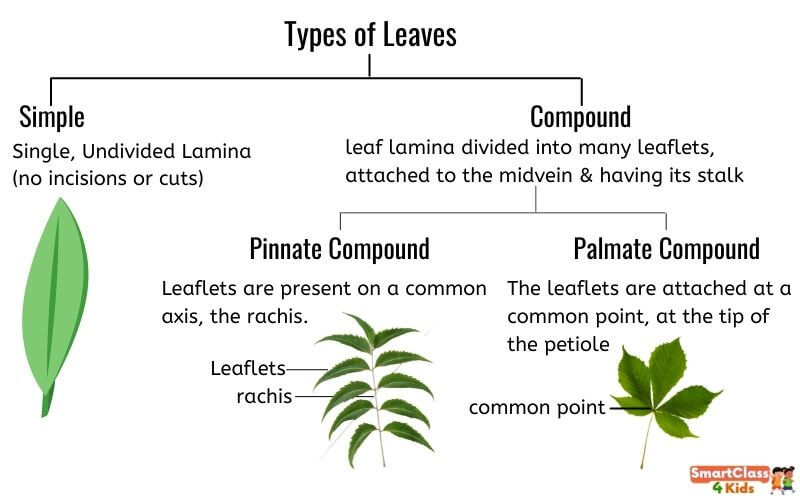
Different types of leaves are described below:
a) Simple leaf: When a leaf's lamina is complete or when the incisions do not touch the midrib, it is considered to be simple.
b) Compound Leaf: The leaf is considered compound when the incisions of the lamina go up to the midrib, breaking it into a number of leaflets. There are two types of compound leaves. As in neem, a pinnatelycompound leaf has a number of leaflets on a common axis, the rachis, which represents the leaf's midrib. The leaflets of palmately compound leaves are joined at a common point, the tip of the petiole, as seen in the silk cotton plant.
In both simple and compound leaves, a bud can be found in the axil of the petiole, but not in the axil of the leaflets.
Phyllotaxy:
The pattern of leaf arrangement on the stem or branch is known as Phyllotaxy. There are three varieties of this: alternate, opposite, and whorled. In alternate phyllotaxy, a single leaf emerges alternately at each node, as seen in China rose, mustard, and sunflower plants. As in Calotropis and guava plants, a pair of leaves emerges at each node and lies opposite each other. This is the opposite phyllotaxy. It is called whorled when more than two leaves emerge from a node and form a whorl, as in Alstonia.

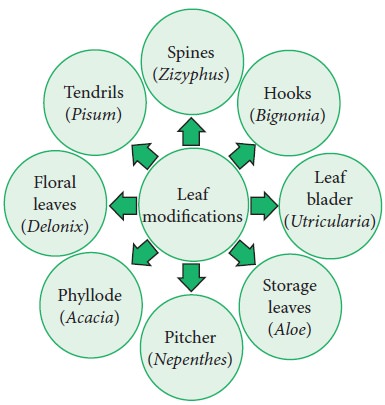
Modifications of a leaf:
Additional to photosynthesis, leaves are frequently adapted to fulfill other purposes. They are transformed into tendrils in peas for climbing and spines in cacti for defence. Onion and garlic leaves have fleshy leaves that store nourishment. The leaves of some plants, such as Australian acacia, are tiny and short-lived. These plants' petioles swell, turn green, and synthesize nourishment. Certain insectivorous plants, such as pitcher plants and venus fly traps, have modified leaves as well.
The Inflorescence
Inflorescence
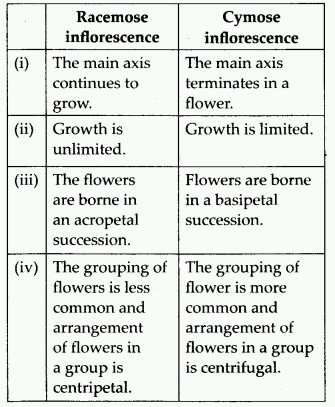
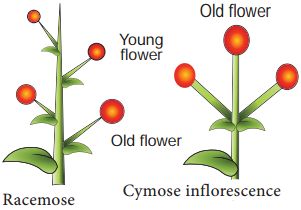
The inflorescence is the arrangement of flowers along the floral axis. A flower is a modified shoot in which the apical meristem of the shoot transforms into the floral meristem. The axis becomes condensed as the internodes do not lengthen. Instead of leaves, the apex produces several types of floral appendages at successive nodes. The transformation of a shoot tip into bloom is always solitary. Two basic forms of inflorescences are described based on whether the apex develops into a flower or continues to grow:
1. Racemose. The primary axis of racemose inflorescences continues to grow, but the flowers are carried laterally in acropetal succession. Younger flowers are present at the tip while older flowers are arranged at the base of this inflorescence. Example: Mustard, Snapdragon, Gulmohar, Wheat, Barley, Parsley.
2. Cymose: The primary axis of a cymose inflorescence terminates in a flower, limiting its expansion.The flowers are borne in a basipetal order. Younger flowers are present at the base of the inflorescence, while older flowers are present at the top. The cymose inflorescence is present in plants like Solanum nigrum, Drosera, Begonia, Ranunculus, Jasmine, Calotropis, etc.
The Flower
The Flower
In angiosperms, the flower is the reproductive unit. It's designed to be used for sexual reproduction. On the swollen end of the stalk or pedicel, called the thalamus or receptacle, a typical flower has four different kinds of whorls placed sequentially. Calyx, corolla, androecium, and gynoecium are the four flower parts. Androecium and gynoecium are reproductive organs, while calyx and corolla are accessory organs. The calyx and corolla of some flowers, such as lilies, are not distinct and are referred to as perianth. A flower is bisexual if it has both androecium and gynoecium. Unisexual flowers are those that have only stamens or carpels.
The flower can be actinomorphic (radial symmetry) or zygomorphic (bilateral symmetry). Flowers that have multiple lines of symmetry (like a starfish) are radially symmetrical, also called actinomorphic. Flowers with only a single line of symmetry are bilaterally symmetrical, also called zygomorphic. Actinomorphic flowers, such as mustard, datura, and chilli, can be divided into two equal radial halves in any radial plane running through the centre. It is zygomorphic when it can be separated into two comparable halves only in one vertical plane, as in pea, Gulmohar, bean, and Cassia. If a flower, like canna, cannot be divided into two comparable halves by any vertical plane going through the centre, it is asymmetric (irregular).

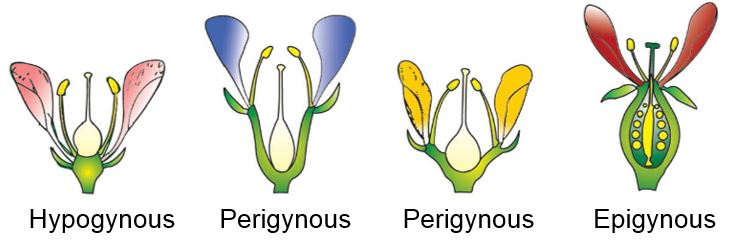
Trimerous, tetramerous, and pentamerous flowers have floral appendages in multiples of 3, 4, and 5, respectively. Bracteate flowers have a shortened leaf at the base of the pedicel, whereas ebracteate flowers do not have one. Flowers are classified as hypogynous, perigynous, or epigynous based on the position of the calyx, corolla, and androecium in relation to the ovary on the thalamus. The gynoecium is at the top of the hypogynous flower, with the other sections below it. Such flowers, such as mustard, China rose, and brinjal are said to have superior ovaries. If the gynoecium is in the center and the other parts of the flower are virtually at the same level as the thalamus rim, the flower is said to be perigynous.Plum, rose, and peach, for example, have a half inferior ovary. The thalamus margin develops upward in epigynous flowers, entirely enclosing and fusing with the ovary; the other parts of the flower ascend above the ovary. As a result, the ovary is reported to be inferior in guava and cucumber flowers, as well as sunflower ray florets.
Parts of a flower include:

Calyx: The calyx is the flower's outermost whorl, and the sepals are its members. Sepals are green, leaf-like structures that protect the flower at the bud stage. The calyx can be gamosepalous (sepals together) or polysepalous (sepals apart) (sepals free).
Corolla: Petals make up the corolla. Petals are typically vividly coloured to attract pollinating insects. Corollas can be gamopetalous (petals joined) or polypetalous, just as the calyx (petals free). Plants have a wide range of corolla shapes and colours. Tubular, bell-shaped, funnel-shaped, or wheel-shaped corollas are all possible.
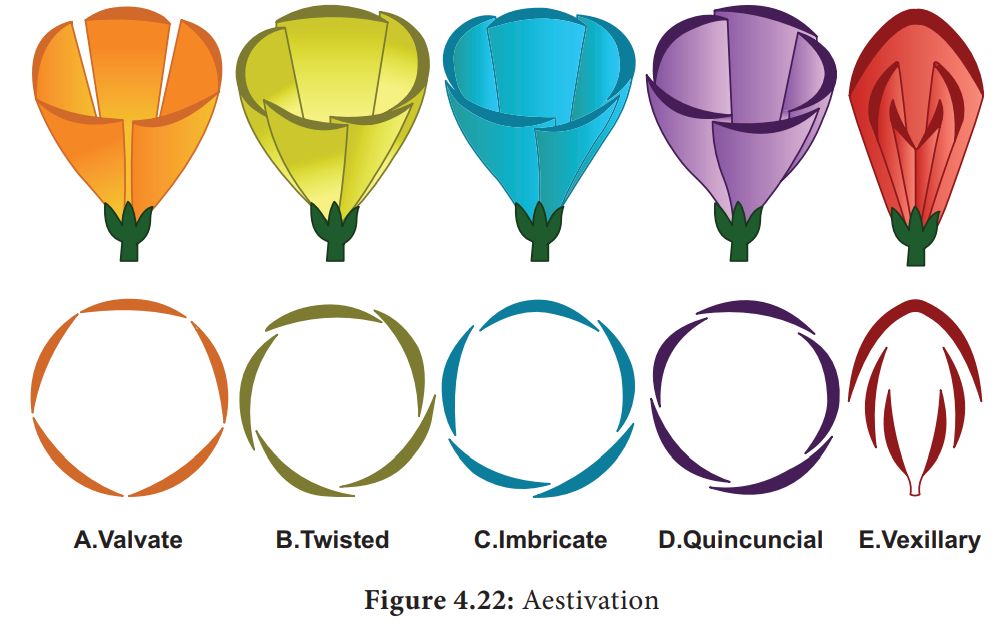
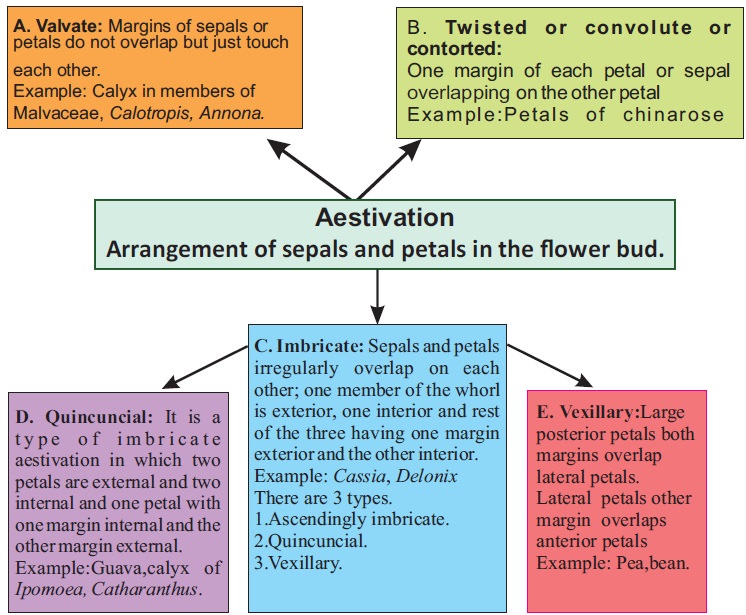
Aestivation refers to the way sepals or petals in a floral bud are arranged in relation to the other members of the same whorl. Valvate, twisted, imbricate, and vexillary aestivation are the most common kinds. Valvate describes a whorl in which the sepals or petals only touch at the margin, without overlapping, as in Calotropis. Twisted means that one appendage's edge overlaps the next and so on, as in the china rose, lady's finger, and cotton. Imbricate aestivation occurs when the borders of sepals or petals overlap but not in any specific direction, as in Cassia and Gulmohar.The largest (standard) petal overlaps the two lateral petals (wings), which overlap the two smallest anterior petals (keel) in pea and bean flowers; this type of aestivation is known as vexillary or papilionaceous.
Androecium: Stamens make up the androecium. A stalk or filament and an anther make up each stamen, which represents the male reproductive organ. Each anther is normally bilobed, with two pollen-sac chambers in each lobe. Pollen grains are manufactured in pollen sacs. The staminode is a sterile stamen. Flower stamens can be joined to other components of the flower, such as petals, or to each other. Stamens are epipetalous when attached to the petals, as in brinjal, or epiphyllous when attached to the perianth, as in lily flowers. A flower's stamens can either remain free (polyandrous) or be joined to varying degrees.The stamens can be grouped into a single bunch or bundle (monoadelphous), two bundles (diadelphous), or more than two bundles (polyadelphous) as in China rose. Within a flower, filament length can vary, as in the case of Salvia and mustard.
Gynoecium: The female reproductive component of the flower is the gynoecium, which is made up of one or more carpels. The stigma, style, and ovary are the three sections of a carpel. The expanded basal portion of the ovary is where the elongated tube, the style, is located. The ovary is linked to the stigma by the style. The stigma is the receptive surface for pollen grains and is usually found at the tip of the style. One or more ovules are linked to a flattened, cushion-like placenta in each ovary. When more than one carpel is present, it is referred to as apocarpous as in lotus and rose. When carpels fuse, as in mustard and tomato, they are called syncarpous.The ovules develop into seeds after fertilization, and the ovary matures into a fruit.
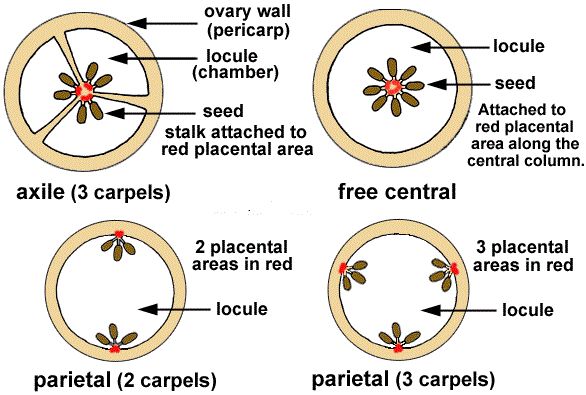
Placentation is defined as the placement of ovules within the ovary. There are several varieties of placentation: marginal, axile, parietal, basal, central, and free central. The placenta forms a ridge along the ventral suture of the ovary in marginal placentation, and the ovules are carried in two rows on this ridge, as in pea. The placenta is considered to be axile when the placenta is axial and the ovules are linked to it in a multilocular ovary, as in the China rose, tomato, and lemon. The ovules develop on the inner wall of the ovary or on the peripheral region in parietal placentation. In mustard and Argemone, the ovary is one-chambered but becomes two-chambered due to the creation of the false septum.The placentation is called free central when the ovules are borne on the central axis and the septa are missing, as in Dianthus and Primrose. As in sunflower and marigold, the placenta develops at the base of the ovary and is linked to a single ovule.
The Fruit
The Fruit
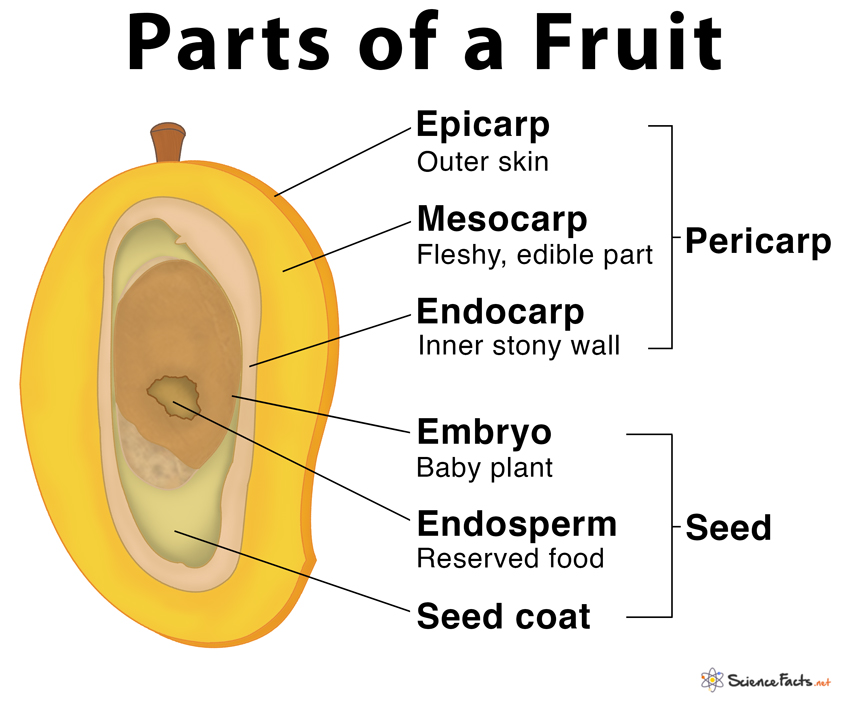
Fruit is a seed-bearing structure generated from the ovary after flowering in flowering plants. Fruits are the vehicles via which flowering plants spread their seeds. Fruit is a distinguishing feature of flowering plants. A ripened that has matured following fertilization is the fruit.
A parthenocarpic fruit is one that is generated without the ovary being fertilized like a banana, cucumber, grape, orange. Etc. A wall or pericarp surrounds the seeds in most fruits. The pericarp might be meaty or dry. The outer epicarp, middle mesocarp, and inner endocarp are formed when the pericarp becomes thick and fleshy. The fruit is called a drupe in mango and coconut. They are single-seeded and develop from monocarpellary superior ovaries.The pericarp of mango is divided into three layers: an exterior thin epicarp, a middle fleshy edible mesocarp, and an inner stony hard endocarp. The mesocarp of coconut, which is also a drupe, is fibrous.
Fruits can be broadly classified into:
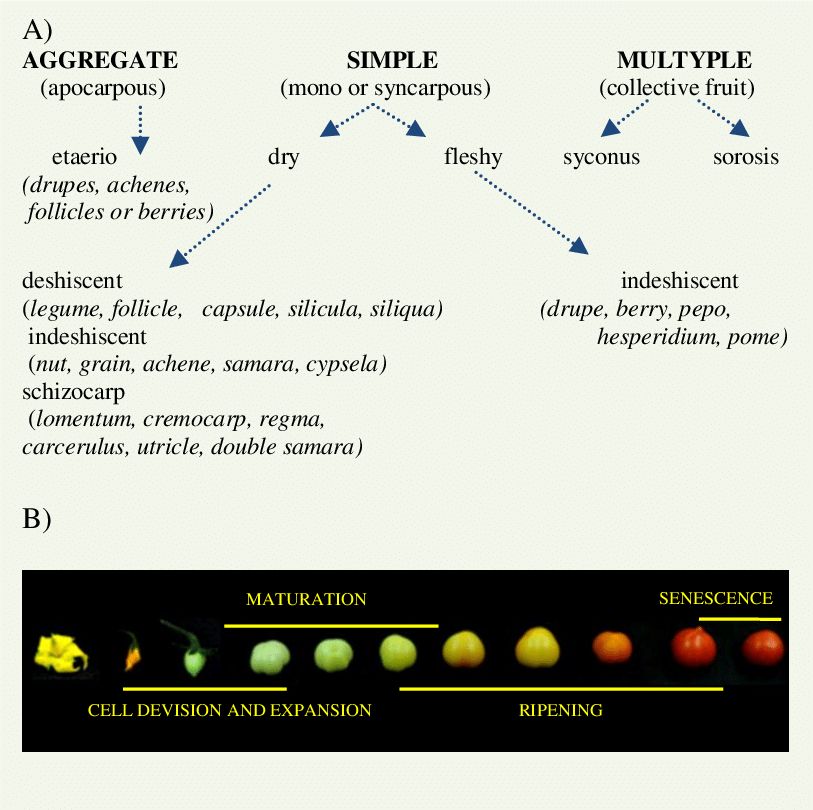
a. Simple Fruit: It is an indehiscent fruit derived from a single ovary having one or many seeds within a fleshy wall or pericarp: e.g. grape; tomato; cranberry.A simple fruit is further classified as dry or fleshy.
b. Aggregate fruit: Itdevelops from a flower having a polycarpellary apocarpous pistil. Each ovary develops into single simple fruit. Examples are blackberries and raspberries where each fleshy lobe is actually an individual fruit joined at their bases.
c. Multiple fruits: They are also called collective fruits. In this, the fruiting bodies are formed from a cluster of flowers, the inflorescence. Each flower in the inflorescence produces a fruit, but these mature into a single mass. After flowering, the mass is called an infructescence.Examples are, Pineapple and Mulberry.
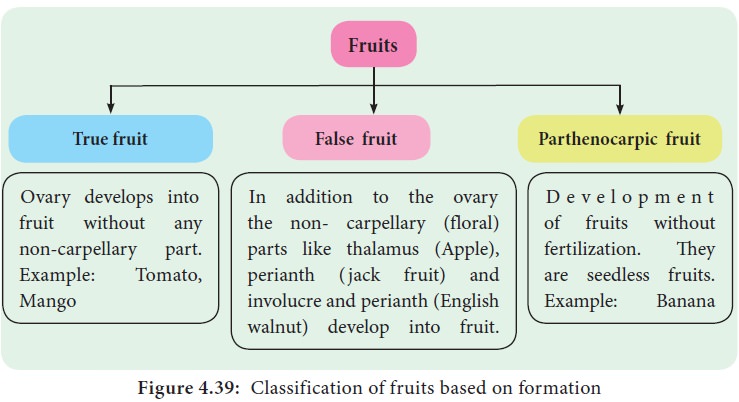
Fruits are also classified on the basis of formation as True fruits, False fruits, and Parthenocarpic fruits.
The Seed
The Seed
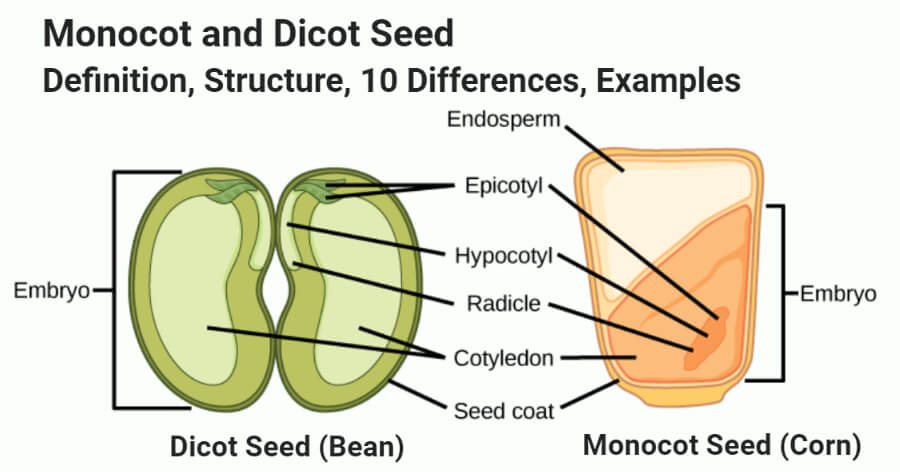
After fertilization, the ovules mature into seeds. A seed consists of two parts: a seed coat and an embryo. A radicle, an embryonal axis, and one (as in wheat, maize) or two cotyledons make up the embryo (as in gram and pea).
Structure of a Dicotyledonous Seed:
The seed coat is a seed's outermost coating. The outer testa and the inner tegmen are the two layers of the seed coat. The hilum is a scar on the seed coat that was used to adhere the developing seeds to the fruit.The micropyle is a tiny pore located above the hilum. The embryo, which consists of an embryonal axis and two cotyledons, is contained within the seed coat. The cotyledons are usually plump and rich in food reserves. The radicle and the plumule are located at the two ends of the embryonal axis. The endosperm developed as a result of multiple fertilization in some seeds, such as castor, is a food-storing tissue and is referred to as endospermic seeds.The endosperm is absent in mature seeds of plants such as beans, gram, and pea, and these seeds are referred to as non-endospermic.
Structure of Monocotyledonous Seed:
Monocotyledonous seeds are usually endospermic, but some, such as those found in orchids, are not. The seed coat of cereal seeds, such as maize, is membranous and usually united with the fruit wall. The endosperm is a thick substance that stores food. A protein layer termed aleurone separates the embryo from the endosperm's outer covering. The embryo is tiny and rests in a groove on one end of the endosperm. It consists of a scutellum, a big shield-shaped cotyledon, and a short axis with a plumule and radicle. Coleoptile and coleorhiza are sheaths that surround the plumule and radicle, respectively.
Semi-Technical Description of a Typical Flowering Plant
Semi-Technical description of a typical flowering plant
A flowering plant is described using morphological characteristics.Vegetative elements such as roots, stems, and leaves are covered first in a semi-technical description of the flowering plant's behavior, followed by floral characters such as inflorescence and flower components.
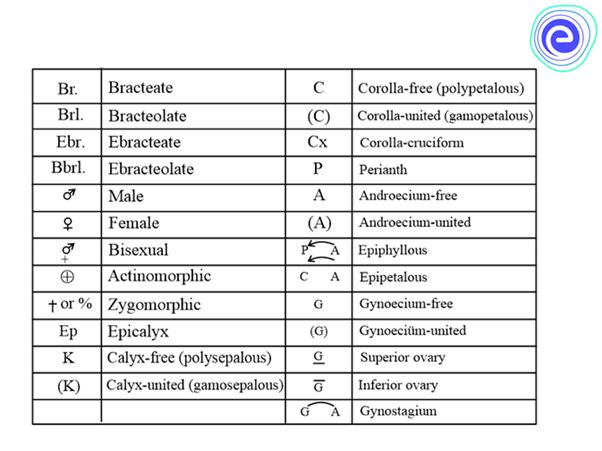
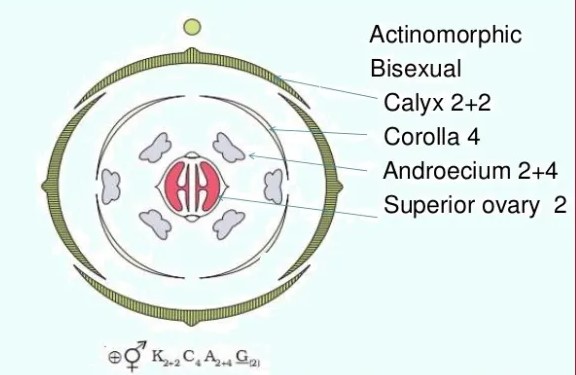
Various symbols in a floral formula are given above. The description must be concise, written in plain, scientific language, and given in a logical order. The plant's habitatand vegetative characteristics (roots, stem, and leaves) are detailed first, followed by floral characters (inflorescence and flower components). A floral diagram is a visual representation of the structure of a flower. It depicts the number, arrangement, and fusion of floral organs. The flower's various components are represented by their respective symbols. Floral diagrams can be used to identify flowers or to better understand angiosperm evolution.The floral formula is a traditional way of describing a flower's structure. It designates different components of the flower with a combination of capital letters and symbols. A floral diagram and a floral formula are offered after discussing various elements of the plant. A floral diagram of a mustard plant is given below.
Some important families
Description of some important families
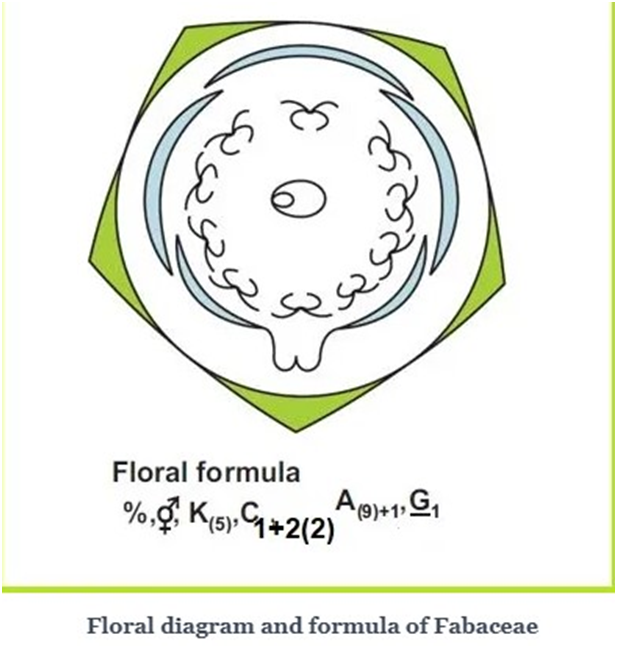
A. FABACEAE
It is also called Leguminosae, It consists of legumes such as the pea, or bean family, and is a large and agriculturally important family of flowering plants.
Vegetative Characters
Root: Dicotyledons, taproot with root nodules.
Stem: Erect or climber; Fabaceae includes shrubs, herbs, trees and majorly climbers.
Leaves: Petiolate, pinnately compound or simple; pulvinus leaf base, stipulate; reticulate venation.
Floral Characters
Inflorescence: Racemose.
Flower: Complete, bisexual, zygomorphic, hypogynous, bracteate/ ebracteate.
Calyx: Five sepals, gamosepalous; valvate or imbricate aestivation.
Corolla: Five petals, polypetalous, papilionaceous, vexillary aestivation.
Androecium: Ten stamens (9+1), diadelphous, anther dithecous.
Gynoecium: Superior ovary, monocarpellary, unilocular, single, short -style and flat, hairy-stigma.
Fruit: Legume.
Seed: One or more, non-endospermic.
Economic Importance
Many plants belonging to this family are economically useful. A few of them are listed below:
The plants of this family are unique and have root nodules that contain nitrogen-fixing symbiotic bacteria, capable of transforming atmospheric nitrogen into fixed nitrogen or ammonia. Pulses like gram, moong, and soya bean are the main source of food. The Mulethiplant is known for its medicinal value. Soya bean and groundnuts are used to extract oil that is used for cooking. Sunn hemp is the source
B. SOLANACEAE
It is a family of flowering plants that ranges from annual and perennial herbs to vines, epiphytes, shrubs, and trees, and includes a number of agricultural crops, medicinal plants, spices, weeds, and ornamentals.
Vegetative Characters
Root System: Taproot system.
Stem: Erect or climber; Solanaceae includes herbs, shrubs, small trees, and climbers.
Leaves: Alternate, simple or pinnately compound (rarely); exstipulate; reticulate venation.
Floral characters
Inflorescence: Racemose- terminal or axillary raceme; Cymose- solitary in Solanum.
Flower: Complete, bisexual, actinomorphic, hypogynous.
Calyx: Five sepals, gamosepalous; valvate aestivation.
Corolla: Five petals, gamopetalous, valvate aestivation.
Androecium: Five stamens, epipetalous; anthers basifixed.
Gynoecium: Syncarpous, bicarpellary, bilocular, superior ovary, axile placentation.
Fruit: Berry/ capsule.
Seed: Numerous, endospermic.
Economic Importance
These are an important source of food. E.g. tomato, brinjal and potato. These are important sources of spices. E.g. chilly. The leaves of Nicotiana tabacum are a major source of tobacco.These are also used as ornamental plants. E.g. petunia. Plants such as Belladonna and Ashwagandha are also used as medicinal plants.

C. LILIACEAE
Liliaceae is the family of around 2500 species of perennial, herbaceous monocots. It is also known as the ‘lily family’.
Vegetative Characters
Root: Fibrous root system.
Stem: Erect; Liliaceae includes perennial herbs which propagate through bulbs or rhizomes.
Leaves: Alternate, simple; exstipulate; parallel venation.
Floral characters
Inflorescence: Cymose- solitary; umbellate clusters.
Flower: Complete, bisexual, actinomorphic; hypogynous, perianth present.
Perianth: Indistinctive sepal and petal; six tepals (3+3), often united tepals; valvate aestivation.
Androecium: Six stamens in two whorls (3+3).
Gynoecium: Syncarpous, tricarpellary, trilocular, superior ovary with axile placentation.
Fruit: Mostly Capsule and sometimes berry.
Seed: Endospermic seeds.
Economic Importance
The plants belonging to the Liliaceae family are a source of Medicine -Aloe vera, Smilax and Colchicine. They are also used as ornamental plants -Lilium, tulips, Gloriosa, and Ruscus. They are also exploited as a source of food (or) Vegetables-Asparagus.Bulbs of Allium cepa and the roots of various species of Smilax are used as flavouring agents.

The Tissues
Chapter 6
Anatomy of flowering plants
Both plants and animals have structural similarities and differences in their outward appearance that can be easily viewed. Furthermore, the interior structure of living creatures shares certain characteristics. The internal structure and organization of higher plants are discussed in this chapter. Anatomy is the study of a plant's interior structure. Cells are the basic unit of plants, and cells are organized into tissues, which are then organized into organs. Different organs of a plant have different internal structures. Monocots and dicots are considered to be physically distinct within angiosperms. Internal structures also exhibit environmental adaptations.
Tissues
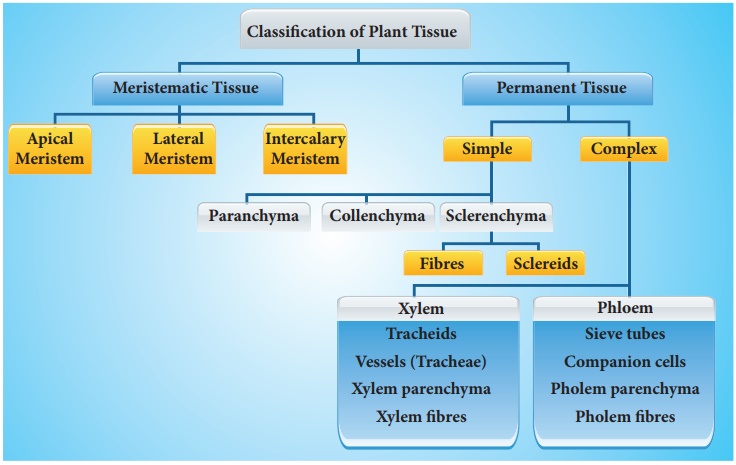
A tissue is a collection of cells that share the same origin and typically execute the same function. Different types of tissues make up a plant. Tissues are divided into two categories: meristematic and permanent tissues, depending on whether or not the cells forming them are capable of dividing.
A. Meristematic Tissues
Meristematic tissue is a type of plant tissue that can divide continuously throughout its existence. Plant growth is generally restricted to meristems, which are specialized regions of active cell division. Meristems come in several forms in plants. Primary meristems include apical and intercalary meristems, which arise early in a plant's life and contribute to the production of the primary plant body.
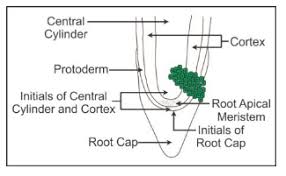
Apical meristems are meristems that form at the terminals of roots and shoots and create primary tissues. The root apical meristem is found at the root's tip, while the shoot apical meristem is found at the far end of the stem axis. The axillary bud is formed by cells left over from the shoot apical meristem after the production of leaves and stem elongation. These buds can develop a branch or a flower and can be found in the axils of leaves.
Intercalary meristem is a type of meristem that arises between mature tissues. They are present in grasses and restore sections of the plant that have been eaten by grazing herbivores. They are found in the leaves and internodes. These contribute to the internode's lengthening. It is also present in monocots, and pine trees. It contributes to the plant's height.
The secondary or lateral meristem is found in the mature portions of roots and shoots of many plants, especially those that generate woody axes and develop later than the primary meristem. They are meristems that are cylindrical. Lateral meristems include the fascicular vascular cambium, interfascicular cambium, and cork-cambium. The secondary tissues are produced by these cells. Following cell division in both primary and secondary meristems, newly produced cells become architecturally and functionally specialized, and their ability to divide is lost. Permanent or mature cells are the cells that make up the permanent tissues. Specific parts of the apical meristem produce dermal tissues, ground tissues, and vascular tissues during the creation of the main plant body.

B. Permanent Tissues:
Permanent tissues are made up of mature cells that have lost their ability to divide and have taken on a permanent shape, size, and function as a result of meristematic tissue development and differentiation. These tissues' cells can be alive or dead, thin-walled or thick-walled. Permanent tissue cells do not usually divide any further. They are divided into simple tissues and complex tissues.
Simple tissues
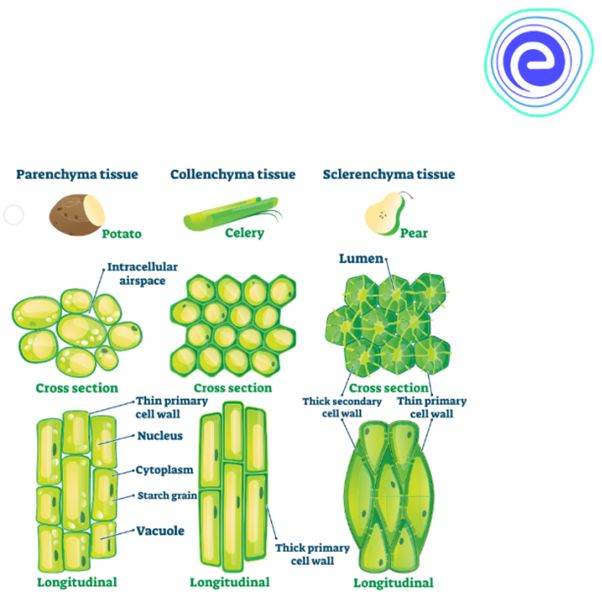
are permanent tissues with all cells having the same structure and function.Only one sort of cell makes up a basic tissue. Plants have parenchyma, collenchyma, and sclerenchyma as simple tissues. The parenchyma cells form a majority of the living cells in the plant. The parenchyma's cells are usually isodiametric. They come in a variety of shapes, including spherical, oval, round, polygonal, and elongated. Their cellulose-based walls are quite thin. They might be tightly packed or with minor intercellular gaps. Photosynthesis, storage, and secretion are all processes performed by the parenchyma. In most dicotyledonous plants, the collenchyma is found in layers beneath the epidermis. It can be found as a single layer or in patches. It is made up of cells that have thickened at the corners due to cellulose, hemicellulose, and pectin deposition.Collenchymatous cells can be oval, round, or polygonal, and chloroplasts are common. When chloroplasts are present, these cells digest food. There are no intercellular spaces. They offer mechanical support to the plant's growth portions, such as the young stem and leaf petiole. Long, narrow cells with thick, lignified cell walls and a few or many pits make up sclerenchyma. They are frequently devoid of protoplasts and lifeless. Sclerenchyma can be classified as fibers or sclereids based on differentforms, structures, origins, and development. Fibers are thick-walled, elongated, cells that can be found in groups throughout the plant. Sclereids are dead cells that are spherical, oval, or cylindrical in shape and have very thin cavities (lumen).These are often found in nut fruit walls, fruit pulp such as guava, pear, and sapota, legume seed coats, and tea leaves. Organs are supported mechanically by sclerenchyma.
Complex tissues
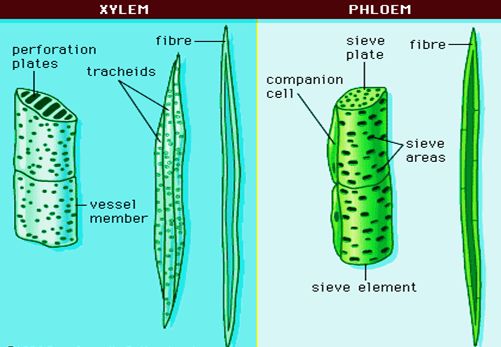
are permanent tissues that contain a variety of cell types.They are made up of multiple cell types that work together as a unit. Plants have complex tissues called xylem and phloem. From the roots to the stem and leaves, the xylem serves as a conduit for water and minerals. It also gives the plant parts mechanical strength. Tracheids, vessels, xylem fibres, and xylem parenchyma are the four types of elements that make up the xylem. The xylem of gymnosperms is devoid of vessels. Tracheids are long, tube-like cells with lignified walls and tapering ends. These have no protoplasm and are dead. The inner layers of the cell walls feature different types of thickenings. Tracheids and vessels are the principal water transportation elements in blooming plants.A vessel is a long cylindrical tube-like structure composed of multiple vessel parts, each with lignified walls and a big central chamber. The vascular cells are also protoplasm-free. Members of the vessel are linked through perforations in their common walls. Angiosperms are characterized by the presence of vessels. The walls of xylem fibers are thicker and the central lumens are destroyed. These can be septate or aseptate in nature. Xylem parenchyma cells are live, thin-walled cells with cellulose-based cell walls. They store food items such as starch or fat, as well as other compounds such as tannins. The ray parenchymatous cells are responsible for water radial conduction.
Protoxylem and metaxylem are the two forms of primary xylem. Protoxylem refers to the first created primary xylem elements, while metaxylem refers to the later formed primary xylem components. The protoxylem is located at the organ's center (pith), while the metaxylem is located near the organ's perimeter. Endarch is the name for this sort of primary xylem. Protoxylem is found on the periphery of roots, while metaxylem is found in the centre. Exarch is the name for this type of primary xylem configuration. Food supplies are transported via phloem from leaves to other sections of the plant. Sieve tube elements, companion cells, phloem parenchyma, and phloem fibres make up phloem in angiosperms. Albuminous cells and sieve cells are found in Gymnosperms. Sieve tubes and companion cells aren't present.
Sieve tube elements are similar to companion cells in that they are long, tube-like structures that are aligned longitudinally. The sieve plates are formed by perforating their end walls in a sieve-like pattern. A developed sieve element has a big vacuole and peripheral cytoplasm but no nucleus. The nucleus of companion cells controls the actions of sieve tubes. Companion cells are specialized parenchymatous cells that are intimately linked to sieve tube elements. Between their common longitudinal walls, pit fields connect the sieve tube elements and companion cells. The partner cells aid in maintaining the sieve tubes' pressure gradient. Elongated, tapered cylindrical cells with rich cytoplasm and nucleus make up phloem parenchyma.The cellulose cell wall has pores through which plasmodesmata connections between cells can be made. Food and other compounds like as resins, latex, and mucilage are stored in the phloem parenchyma. The majority of monocotyledons lack phloem parenchyma. Sclerenchymatous cells make up phloem fibers. The primary phloem is devoid of them, whereas the secondary phloem has them. These are long and unbranched, with sharp, needle-like apices. Phloem fibers have a very thick cell wall. These fibers lose their protoplasm and die as they mature. Protophloem is the first created primary phloem, which has tiny sieve tubes, and metaphloem is the second formed primary phloem, which has larger sieve tubes.
The Tissue System
The Tissue System
There are three types of tissue systems based on their structure and location.
A. The epidermal tissue system
Epidermal cells, stomata, and epidermal appendages like trichomes and hairs make up the epidermal tissue system, which covers the entire plant body. The epidermis is the major plant body's outermost layer. It is made up of a continuous layer of elongated, compactly packed cells. The epidermis is normally one layer thick. Epidermal cells are parenchymatous, with a big vacuole and a little quantity of cytoplasm along the cell wall. The cuticle, a waxy thick covering on the outside of the epidermis that resists water loss, is commonly present. The roots have no cuticle. The epidermis of leaves has features called stomata. The process of transpiration and gas exchange is regulated by stomata.Guard cells enclose the stomatal pore in each stoma, which are two bean-shaped cells. The guard cells in grasses are formed like a dumbbell. Guard cells have thin outer walls that face away from the stomatal pore and robust interior walls that face the stomatal pore. Guard cells have chloroplasts and control stomata opening and closing. A few epidermal cells near the guard cells can become specialized in shape and size, and these cells are known as subsidiary cells. The Stomatal apparatus is made up of the stomatal orifice, guard cells, and surrounding subsidiary cells. Hairs are found on the epidermis cells.Root hairs are unicellular epidermal elongations that assist absorb water and minerals from the soil. Trichomes are epidermal hairs that grow on the stem. In the shoot system, trichomes are frequently multicellular. They might be soft or stiff, branching or unbranched. They could even be hidden. The trichomes aid in the prevention of water loss by transpiration.
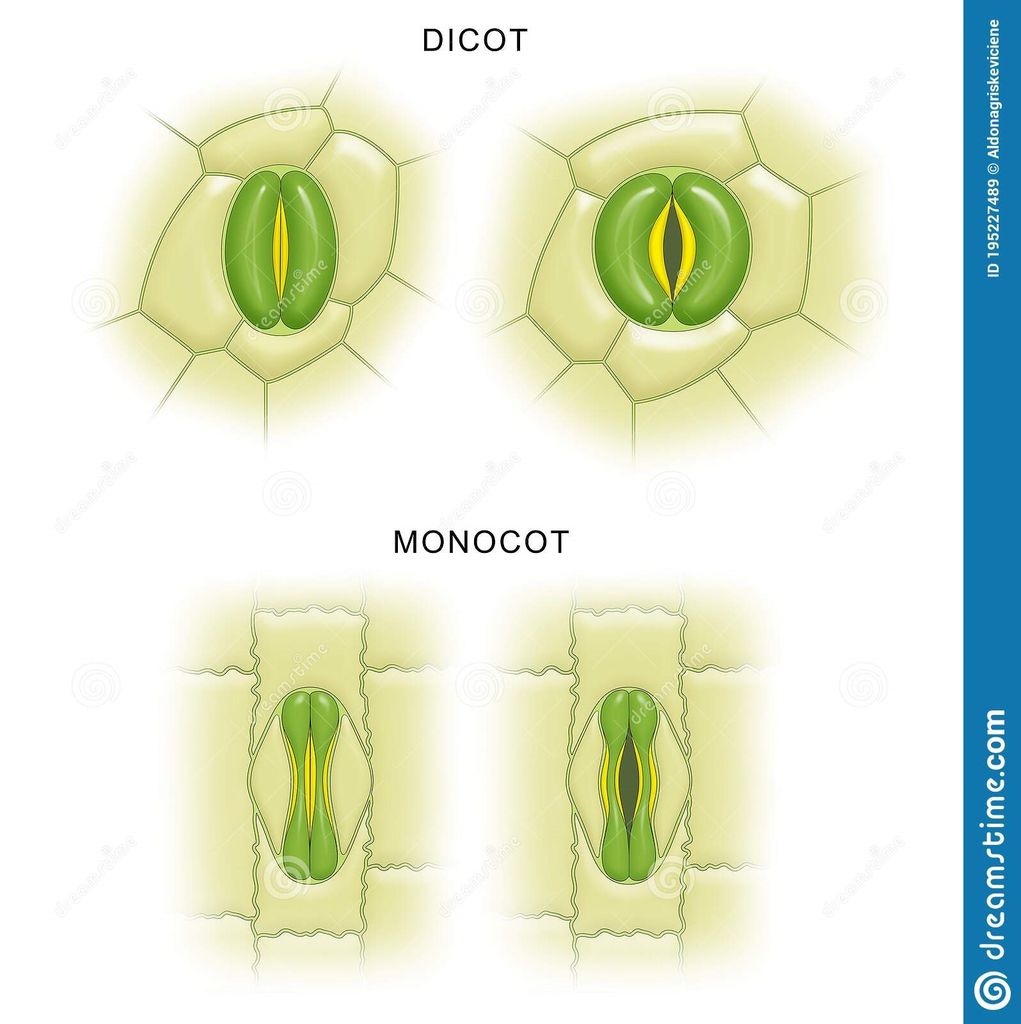
B. The ground tissue system
It is also called the foundational tissue system.The ground tissue is made up of all tissues save the epidermis and vascular bundles. Simple tissues such as parenchyma, collenchyma, and sclerenchyma make up this layer. In the cortex, pericycle, pith, and medullary rays, as well as the primary stems and roots, parenchymatous cells are commonly seen. The mesophyll is the ground tissue in leaves that is made up of thin-walled chloroplast-containing cells. Ground tissue includes parenchyma (photosynthesis in the leaves and storage in the roots), collenchyma (shoot support in areas of active growth), and sclerenchyma (shoot support in areas where growth has ceased) and is the site of photosynthesis, provides a supporting matrix for the vascular tissue, provides structural support for the stem, and helps to store water and sugars, depending on the cell type and location in the plant.
C. The vascular tissue system:
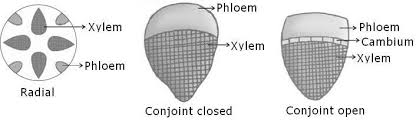
Also called conducting tissue system. The vascular system is made up of two complicated tissues: phloem and xylem. Vascular bundles are made up of the xylem and phloem. Between the phloem and xylem in dicotyledonous stems, there is cambium. Because of the existence of cambium, such vascular bundles have the ability to produce secondary xylem and phloem tissues, and are hence referred to as open vascular bundles. The cambium is absent from the vascular bundles of monocotyledons. As a result, they are referred to as closed since they do not generate additional tissues. Radial arrangement occurs when xylem and phloem within a vascular bundle are placed in an alternative fashion along distinct radii, as in roots.The xylem and phloem are located along the same radius of vascular bundles in conjoint type vascular bundles. In stems and leaves, vascular bundles are common. Phloem is usually exclusively seen on the outer side of the xylem in conjoint vascular bundles.
Anatomy of Dicotyledonous and Monocotyledonous Plants
Anatomy of dicotyledonous and monocotyledonous plants
Transverse sections of the mature zones of roots, stems, and leaves are useful for better understanding the tissue organization of these organs.
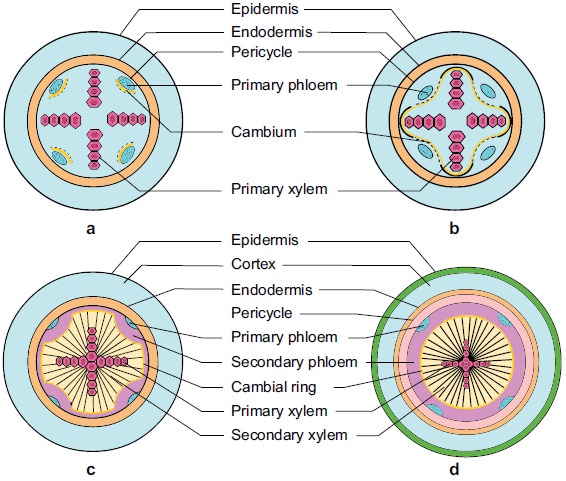
A.Dicotyledonous Root: The interior tissue organization is as follows:
1. Epiblema is the outermost layer in the interior tissue architecture. Epiblema cells emerge as unicellular root hairs on many occasions.
2. The cortex is made up of many layers of parenchymal cells with thin walls and intercellular gaps.
3. Endodermis is the cortex's deepest layer. There are no intercellular gaps in the single layer of barrel-shaped cells.
4. Water-impermeable waxy substance suberin is deposited in the form of Casparian strips on the tangential as well as radial walls of endodermal cells.
5. A few layers of thick-walled parenchymatous cells called pericycle are found next to the endodermis. During secondary growth, these cells initiate the formation of lateral roots and vascular cambium.
6. The pith is small and undetectable.
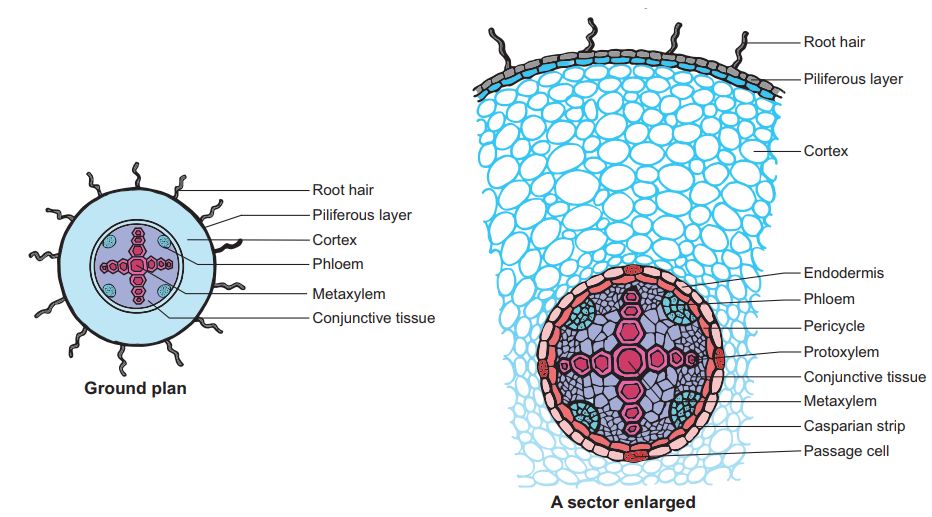
7. Conjuctive tissue is made up of parenchymatous cells that reside between the xylem and the phloem. Two to four xylem and phloem patches are typical. A cambium ring forms later between the xylem and the phloem.
8. The stele is made up of all tissues on the inner side of the endodermis, such as the pericycle, vascular bundles, and pith.
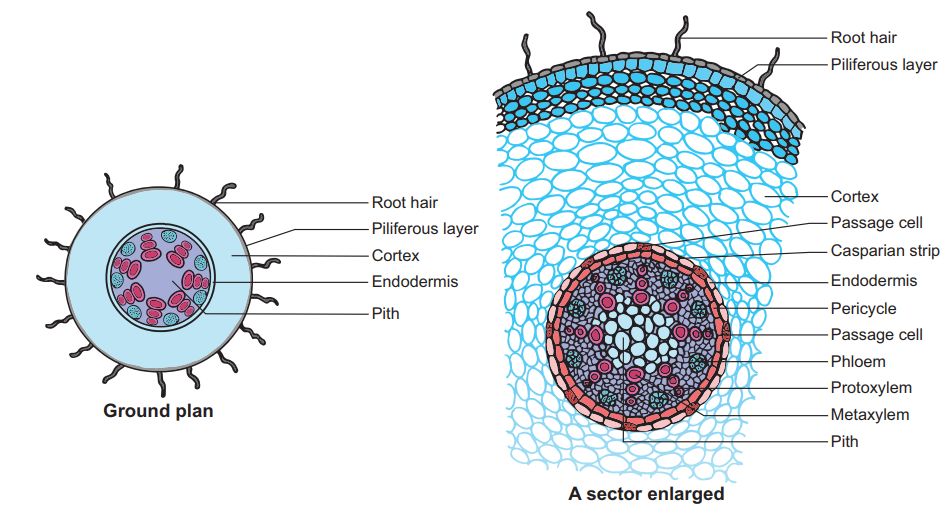
B. Monocotyledonous Root: In many ways, the monocot root's anatomy is identical to that of the dicot root.
1. Epidermis, cortex, endodermis, pericycle, vascular bundles, and pith are all present.
2. Unlike the dicot root, which has fewer xylem bundles, the monocot root frequently has more than six (polyarch) xylem bundles.
3. The pith is thick and developed.
4. There is no secondary development in monocotyledonous roots.
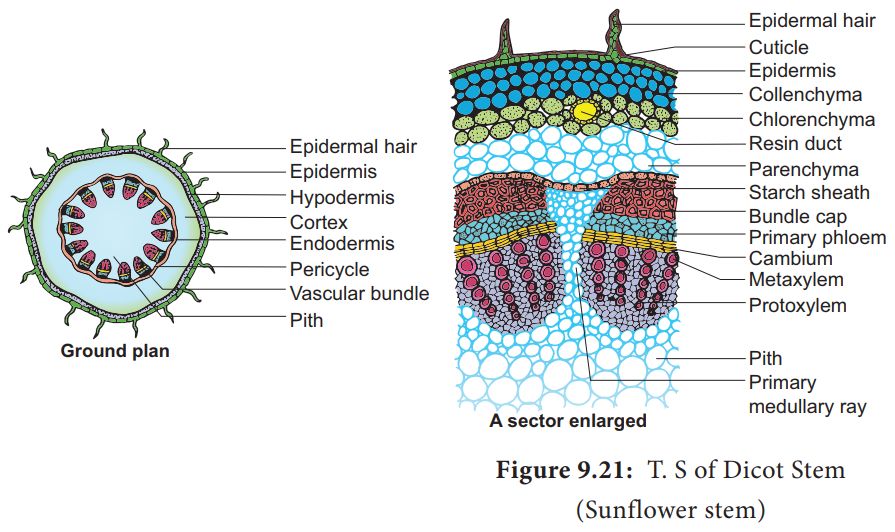
C. Dicotyledonous Stem:
1. The epidermis is the stem's outermost protective covering, as shown in this transverse section of a typical immature dicotyledonous stem.
2. It may have trichomes and a few stomata and is covered in a thin layer of cuticle.
3. The cortex is made up of cells organized in many layers between the epidermis and the pericycle. It is divided into three zones.
4. Just below the epidermis, the outer hypodermis consists of a few layers of collenchymatous cells that offer mechanical support to the embryonic stem.
5. The cortical layers underneath the hypodermis are made up of spherical parenchymatous cells with visible intercellular gaps.
6. The endodermis is the deepest layer andbecause the endodermis cells are densely packed with starch grains, the layer is also known as the starch sheath.
7. Pericycle appears as semi-lunar patches of sclerenchyma on the inner side of the endodermis and above the phloem.
8. Medullary rays are a few layers of radially arranged parenchymatous cells that lie between the vascular bundles.
9. A ring of vascular bundles surrounds the stem of a dicot. Each vascular bundle is joined, open, and has a protoxylem at the end.
10. The pith is made up of a large number of spherical, parenchymatous cells with extensive intercellular gaps that occupy the middle region of the stem.
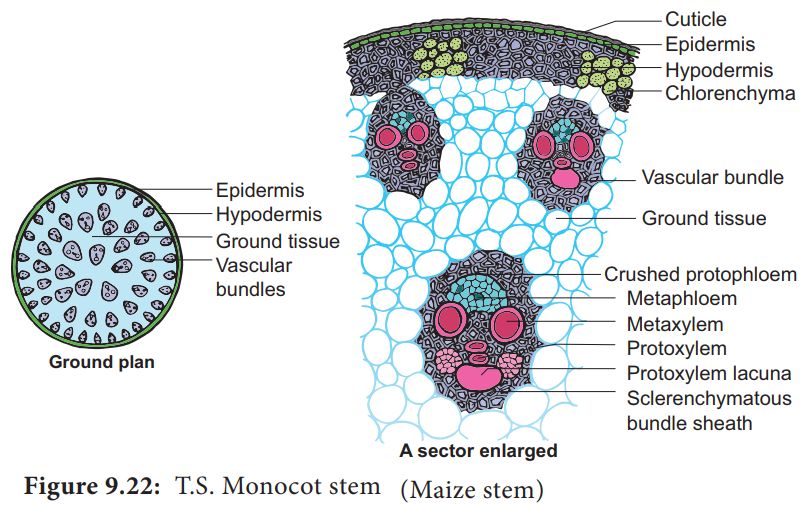
D. Monocotyledonous Stem:
1.A sclerenchymatous hypodermis, numerous distributed vascular bundles, each enclosed by a sclerenchymatous bundle sheath, and a broad, prominent parenchymatous ground tissue characterize the monocot stem.
2. The vascular bundles are joined and closed together. Vascular bundles at the periphery are typically smaller than those in the center.
3. The phloem parenchyma is missing, and there are water-filled holes inside the vascular bundles.
E. Dicotyledonous Leaf:
They are also called Dorsiventral Leaves.
1. The epidermis, mesophyll, and vascular system are visible in a vertical piece of a dorsiventral leaf through the lamina.
2. The epidermis of the leaf has a prominent cuticle that covers both the upper (adaxial epidermis) and bottom (abaxial epidermis).
3. In general, the abaxial epidermis has more stomata than the adaxial epidermis.
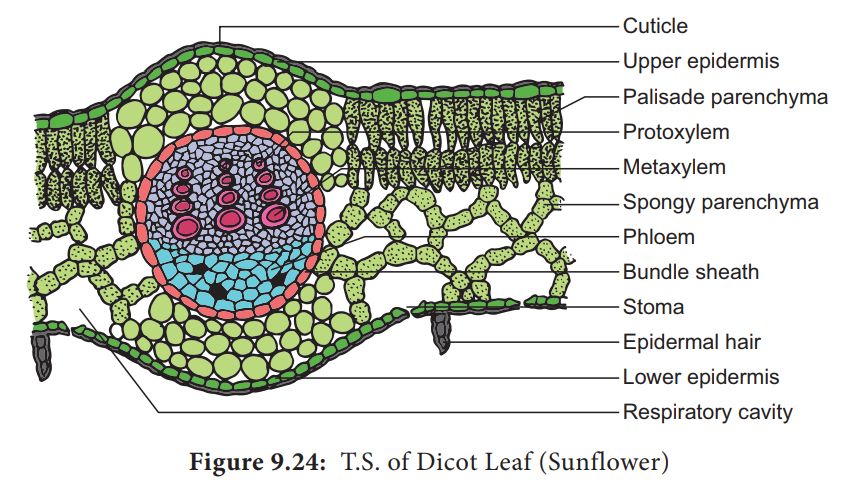
4. Stomata may even be absent in the upper epidermis. The mesophyll is the tissue that lies between the top and lower epidermis.
5. The mesophyll is made up of parenchyma, which contains chloroplasts and performs photosynthesis.
6. The palisade parenchyma and the spongy parenchyma are two types of cells. The elongated cells that make up the adaxial palisade hepatocytes are organized vertically and parallel to each other.
7. The spongy parenchyma, which is oval or spherical and loosely organized, is found underneath the palisade cells and reaches the lower epidermis. Between these cells are various huge gaps and air cavities.
8. Vascular bundles can be found in the veins and midrib of the vascular system. The size of the vascular bundles is determined by the vein size.
9. The veins in the dicot leaves' reticulate venation vary in thickness. A layer of thick-walled bundle sheath cells surrounds the vascular bundles.
F. Monocot Leaf:
They are also known as Isobilateral Leaves.In many ways, the anatomy of the isobilateral leaf is identical to that of the dorsiventral leaf. It demonstrates the following distinguishing characteristics.
1. Stomata are present on both epidermal surfaces in an isobilateral leaf, and the mesophyll is not divided into palisade and spongy parenchyma.
2. Certain adaxial epidermal cells along veins in grasses transform into huge, empty, colorless cells called Bulliform cells.
3. The leaf surface is exposed when the bulliform cells in the leaves have absorbed water and are turgid. They curl the leaves inwards to reduce water loss when they are flaccid due to water stress.
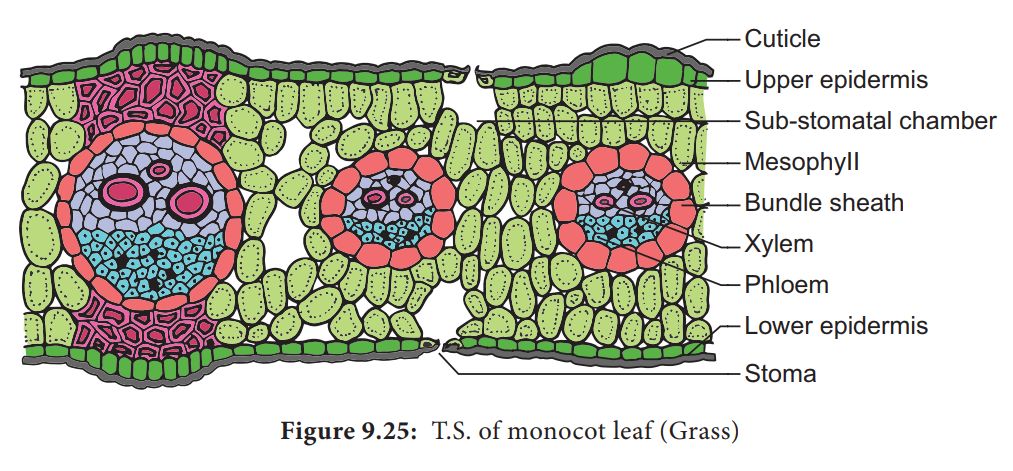
4. In vertical sections of monocot leaves, the parallel venation is represented in the near comparable diameters of vascular bundles.
Secondary Growth
Secondary Growth
Primary growth refers to the lengthening of roots and stems with the help of the apical meristem. Most dicotyledonous plants expand in girth in addition to main growth. This is referred to as secondary growth. The two lateral meristems, vascular cambium and cork cambium, are engaged in secondary growth.
A. Vascular Cambium:
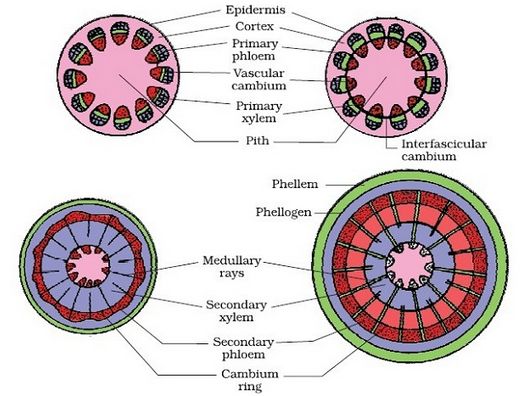
Vascular cambium is the meristematic layer responsible for cutting off vascular tissues – xylem and phloem. It appears in patches as a single layer between the xylem and phloem in the young stem. It eventually forms a complete ring.
i) Formation of cambial ring:
The intrafascicular cambium is the cambium cells found between the primary xylem and primary phloem in dicot stems. The interfascicular cambium is formed when the cells of medullary rays next to this intrafascicular cambium become meristematic. As a result, a continuous ring of cambium forms.
ii) Activity of the cambial ring:
The cambial ring becomes active and starts cutting off new cells from both the inside and outside. Secondary xylem develops from cells cut off towards the pith, while secondary phloem develops from cells cut off towards the periphery. The inner side of the cambium is often more active than the outer side. As a result, secondary xylem is formed in greater quantities than the secondary phloem, forming a compact mass. The persistent development and accumulation of secondary xylem gradually crushes the primary and secondary phloems. The principal xylem, on the other hand, is mostly intact at or near the centre.The cambium sometimes produces a narrow band of parenchyma that runs in radial directions through the secondary xylem and secondary phloem. These are the secondary medullary rays.
iii) Springwood and autumn wood:
Cambium activity is influenced by a variety of physiological and environmental variables. Climate conditions in temperate zones are rarely consistent throughout the year. The cambium is particularly active in the spring and produces a significant number of xylary components with larger vessels. Springwood, also known as earlywood, is formed during this season. Autumn wood or latewood is formed when the cambium is less active in the winter and produces fewer xylary components with thin channels. Springwood is lighter in color and denser, whereas fall wood is darker and denser. An annual ring is made up of two types of wood that appear as alternate concentric rings.Annual rings in a cut stem can be used to estimate the age of a tree.
iv) Heartwood and sapwood:
The deposition of organic components such as tannins, resins, oils, gums, aromatic chemicals, and essential oils in the middle or innermost layers of the stem causes the majority of secondary xylem in ancient trees to be dark brown. These compounds make it tough, long-lasting, and resistant to bacteria and insects. Heartwood is a zone made up of dead components with heavily lignified walls. The stem is supported mechanically by the heartwood, which does not conduct water. Sapwood refers to the lighter-colored portion of the secondary xylem. It helps to transport water and minerals from the root to the leaf.
B.Cork Cambium:The outer cortical and epidermal layers break down as the stem grows in girth due to vascular cambium activity, and they must be replaced to create new protective cell layers. As a result, additional meristematic tissue is known as cork cambium or phellogen forms, usually in the cortical region, sooner or later. Phellogen is made up of several layers. It is made up of cells that are tiny, thin-walled, and practically rectangular. Both sides of the cell are cut off by Phellogen. Inner cells differentiate into secondary cortex or phelloderm, whereas exterior cells differentiate into cork or phellem. Due to suberin deposition in the cell wall, the cork is impenetrable to water. Secondary cortical cells are parenchymatous. The periderm is the combination of phellogen, phellem, and phelloderm.
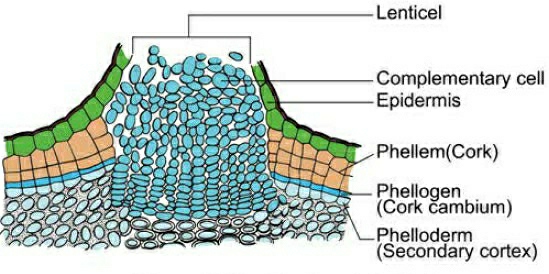
Pressure goes up on the remaining layers peripheral to phellogen due to cork cambium activity, and these layers eventually die and peel off. Secondary phloem is included in the phrase "bark," which is a non-technical term that refers to all tissues outside of the vascular cambium. Periderm and secondary phloem are two forms of tissue that makeup bark. Early or soft bark refers to bark that forms early in the season. Late or hard bark develops toward the end of the season. Instead of cork cells, the phellogen breaks off tightly packed parenchymatous cells on the outer surface in some areas. These parenchymatous cells quickly break the epidermis, generating lenticels, which are lens-shaped holes.Lenticels allow gas exchange between the outside atmosphere and the stem's internal tissue. Most woody trees have these.

C. Secondary Growth in Roots:
The vascular cambium in dicot roots is entirely secondary in origin. It develops from tissue right beneath the phloem bundles, a piece of pericycle tissue, and above the protoxylem, forming a complete and continuous wavy ring that eventually becomes circular. The next events are similar to those outlined before for a dicotyledon stem. Gymnosperm stems and roots also experience secondary growth. Monocotyledons, on the other hand, do not have secondary growth. Secondary growth usually results in a thickening of the root diameter due to the addition of vascular tissue. When cells in the residual procambium and sections of the pericycle begin to make periclinal divisions, secondary growth begins. Periclinal divisions begin only in the pericycle cells opposite the xylem sites. The vascular cambium is formed by the inner layer of cells. The pericycle is the outer layer that is retained. Around the primary xylem, the vascular cambium is continuous. The periclinal division of the vascular cambium continues. If the daughter cells divide towards the core of the root, they become secondary xylem cells, and if they divide towards the outer surface of the root, they become secondary phloem cells.The periderm, which comes from the pericycle and replaces the epidermis, forms an outer protective layer on certain roots. The pericycle restores its meristematic nature and divides periclinallyonce more and thus forms the phellogen or cork cambium. To the outside of the plant, the cork cambium generates phellum cells (cork cells). At maturity, these cells are lifeless. They've been suberized, which makes the cells water-resistant. Phellum cells in cross-section are neatly organized into files. The phelloderm, a mature tissue made up of live cells, is likewise produced by the cork cambium.
Animal Tissues
Chapter 7
STRUCTURAL ORGANISATION IN ANIMALS
In unicellular organisms, a single cell performs all tasks such as digesting, respiration, and reproduction. The same basic duties are carried out by different groups of cells in a well-organized manner in the complex body of multicellular organisms. The body of a basic organism like Hydra is made up of many distinct types of cells, each with thousands of them. The human body is made up of billions of cells that each perform a different purpose. A set of related cells, as well as intercellular chemicals, execute a defined purpose in multicellular creatures. Tissue is the name for this type of arrangement.
There are just four fundamental types of tissues in all complex creatures. To form an organ such as the stomach, lung, heart, or kidney, these tissues are organized in a certain proportion and pattern. When two or more organs interact physically and/or chemically to fulfill a shared function, they create an organ system, such as the digestive system or the respiratory system. Cells, tissues, organs, and organ systems divide up the work and contribute to the body's overall survival through the division of labor.
Animal Tissues
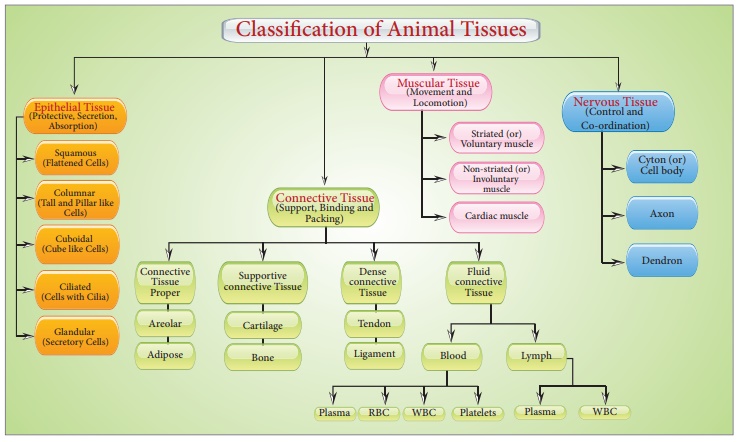
Cells have different structures depending on their purpose. As a result, the tissues differ and can be divided into four categories:
Epithelial Tissue
Epithelial Tissue
Epithelium or epithelia refers to epithelial tissue. This tissue has a free surface that is exposed to either a body fluid or the outside environment and serves as a covering or lining for a body part. The cells are densely packed and there is a little intercellular matrix. Simple epithelium and compound epithelium are two forms of epithelial tissues.
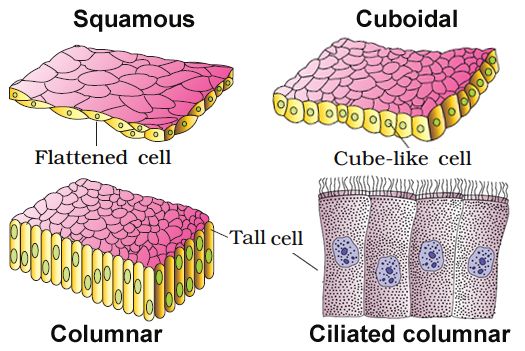
(A) SIMPLE EPITHELIUM: A single layer of cells makes up simple epithelium, which serves as a lining for bodily cavities, ducts, and tubes. Basedon structural modification of the cells, the simple epithelium is further divided into three types. These are
(i) Squamous epithelium: A single thin layer of flattened cells with uneven borders makes up the squamous epithelium. They're present in the walls of blood arteries and the air sacs of the lungs, and they help build diffusion boundaries.
(ii) Cuboidal epithelium: A single layer of cube-like cells makes up the cuboidal epithelium. Its main roles are secretion and absorption, and it is usually found in gland ducts and tubular sections of nephrons in the kidneys. Microvilli are found in the epithelium of the proximal convoluted tubule (PCT) of the nephron in the kidney.
(iii) Columnar epithelium: A single layer of tall and thin cells makes up the columnar epithelium. Their nuclei are near the bottom of their bodies. Microvilli may exist on a free surface. They help with secretion and absorption and are located in the lining of the stomach and intestine.
(iv) Ciliated epithelium: Ciliated epithelium is defined as columnar or cuboidal cells with cilia on their free surfaces. Their job is to transfer particles or mucus over the epithelium in a precise direction. They're mostly found on the inside of hollow organs like the bronchioles and fallopian tubes.
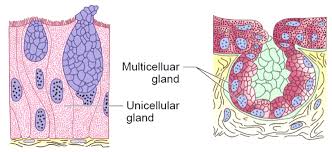
(v) Glandular epithelium: The glandular epithelium is formed when some columnar or cuboidal cells become specialized for secretion.They are divided into two types: unicellular glandular cells (goblet cells of the alimentary canal) and multicellular glandular cells (clusters of cells) (salivary gland). Exocrine and endocrine glands are split into two types based on how they secrete their secretions. Mucus, saliva, earwax, oil, milk, digestive enzymes, and other cell products are secreted by exocrine glands. Ducts or tubes are used to release these products. Endocrine glands, on the other hand, lack ducts. Hormones, which are their products, are secreted straight into the gland's fluid bath.
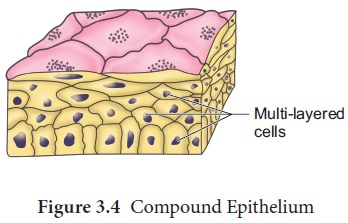
(B) COMPOUND EPITHELIUM: Compound epithelium is made up of multiple layers of cells, therefore it has little function in secretion and absorption. The compound epithelium, like our skin, is made up of two or more cell layers and serves a protective role.Their primary purpose is to protect the body against chemical and mechanical stress. They protect the skin's dry surface, the moist surface of the buccal cavity, the pharynx, the inner lining of the salivary gland, and pancreatic ducts.
The epithelium's cells are bound together by a thin layer of intercellular substance. Specified junctions offer structural and functional linkages between individual cells in practically all animal tissues. In the epithelium and other tissues, there are three types of cell junctions. Tight, adherent, and gap junctions are the three types of junctions. Tight connections prevent chemicals from leaking through a cell's surface.Adhering junctions bind cells by gluing them together. Gap junctions allow cells to communicate with one another by connecting the cytoplasm of neighboring cells, allowing for the rapid movement of ions, small molecules, and occasionally large molecules.Adhering junctions bind cells together by gluing them together.
Connective Tissue
Connective Tissue
In the bodies of sophisticated animals, connective tissues are the most abundant and widely dispersed. Connective tissues get their name from their unique ability to join and support other body tissues and organs. They include soft connective tissues as well as specialized forms such as cartilage, bone, adipose tissue, and blood. Cells in all connective tissues, except blood, secrete collagen or elastin fibers, which are structural proteins. The fibers provide the tissue its strength, elasticity, and flexibility. These cells also release modified polysaccharides, which form a matrix between cells and fibers (ground substance). There are three forms of connective tissue:
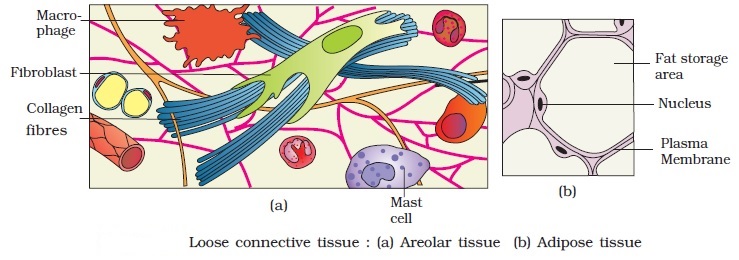
1. Loose connective tissue: Areolar tissue present beneath the skin, is loose connective tissue with cells and fibers loosely distributed in a semi-fluid ground substance. It frequently acts as a support structure for epithelium. Fibroblasts (cells that generate and secrete fibers), macrophages, and mast cells are all present. Another form of loose connective tissue found primarily beneath the skin is adipose tissue. This tissue's cells arespecialized for fat storage. Excess nutrients are converted to lipids and stored in this tissue if they are not utilized right away.
2. Dense connective tissue: The dense connective tissues are densely packed with fibers and fibroblasts. Dense regular and dense irregular tissues have a regular or irregular pattern in their fiber orientation.Collagen fibers are found in rows between several parallel bundles of fibers in dense regular connective tissues. Tendons that connect skeletal muscles to bones and ligaments that connect two bones are examples of this tissue. Fibroblasts and numerous fibers (mainly collagen) are orientated differently in dense irregular connective tissue. The skin contains this tissue.
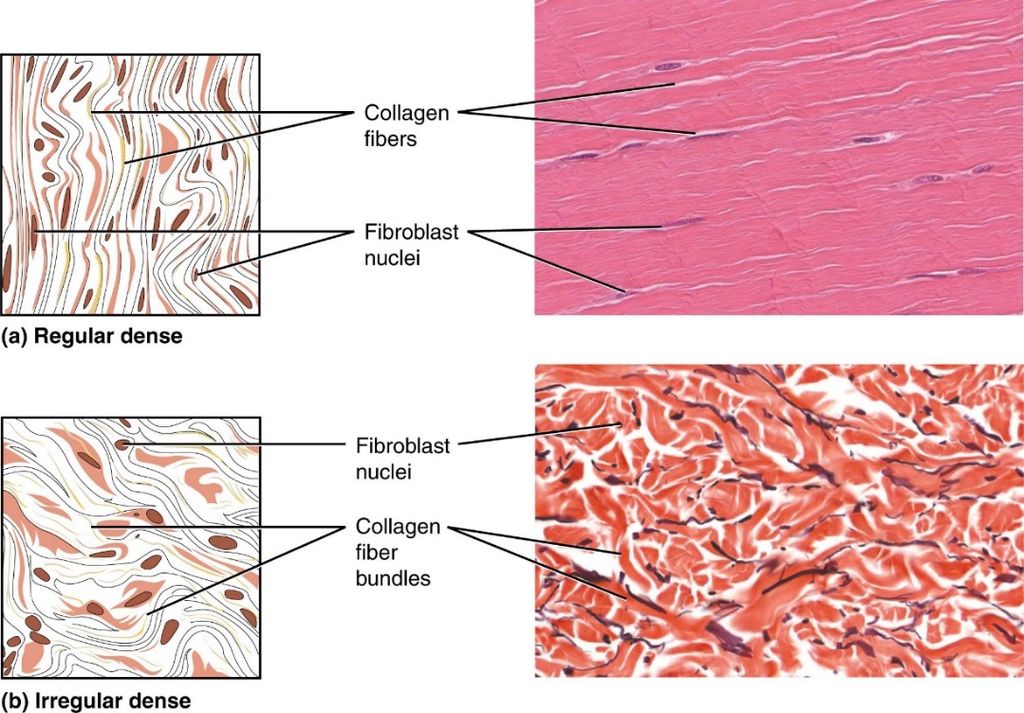
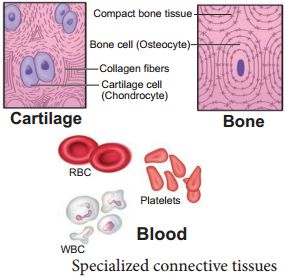
3. Specialised connective tissue: Specialized connective tissues include cartilage, bones, and blood. The cartilage intercellular substance is firm and malleable, and it resists compression. Chondrocytes (tissue cells) are encased in little cavities within the matrix they secrete. In adults, most cartilages in vertebrate embryos are replaced by bones. In adults, cartilage can be found in the tip of the nose, outer ear joints, and between neighboring spinal column bones, limbs, and hands. Bones have a hard, non-pliable ground substance that is rich in calcium salts and collagen fibers, and this is what gives them their strength. It is the major tissue that gives the body its structural framework. Soft tissues and organs are supported and protected by bones.Lacunae are places where bone cells (osteocytes) can be found. The lengthy bones of the legs, for example, serve as weight-bearing structures. They also interact with the skeletal muscles that are linked to them in order to form blood cells in the marrow of some bones. Plasma, red blood cells (RBC), white blood cells (WBC), and platelets make up blood, which is a fluid connective tissue. It is the primary circulating fluid that aids in the transportation of a variety of chemicals.
Muscular Tissue
Muscular Tissue
Many long, cylindrical fibers are organized in parallel arrays in each muscle. Myofibrils are a type of tiny fibril that makes up these fibers. Muscle fibers contract (shorten) in response to stimulus, then relax (lengthen) and uncontract in a coordinated manner. Their actions cause the body to move in order tochange the environment and maintain the postures of the various body parts. The sarcolemma is the muscle cell membrane, and the sarcoplasm is the muscle cell cytoplasm. The excitability of the sarcolemma allows it to conduct electrical impulses that occur during depolarization. Muscles are involved in all of the body's movements in general. Skeletal, smooth, and cardiac muscles are the three types of muscles. Skeletal muscle tissue is tightly bound to the bones. Striated (striped) skeletal muscle fibers are bundled together in a parallel pattern in a typical muscle like the biceps.Several bundles of muscle fibers are encased in a strong connective tissue sheath.Tendons connect skeletal muscles to bones, and they are responsible for all motions of bodily components in relation to one another. Skeletal muscle, unlike smooth and cardiac muscle, is controlled by the user. Shoulder muscles, hamstring muscles, and abdominal muscles are all examples of skeletal muscles. Smooth muscle fibers have no striations and taper at both ends (fusiform). They are held together by cell connections and bundled in a connective tissue sheath. This type of muscle tissue can be found in the walls of internal organs such as the blood vessels, stomach, and intestine. Smooth muscles are referred to as "involuntary" since they cannot be controlled directly. Cardiac muscle tissue is only found in the heart, where it performs coordinated contractions that allowthe heart to pump blood through the circulatory system.The only contractile tissue found in the heart is cardiac muscle tissue. The plasma membranes of heart muscle cells are fused.At some fusion points, communication junctions (intercalated discs) allow cells to contract as a group, meaning that when one cell receives a signal to contract, its neighbors are also stimulated to contract.
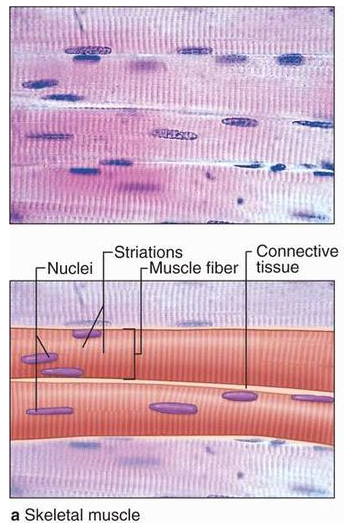
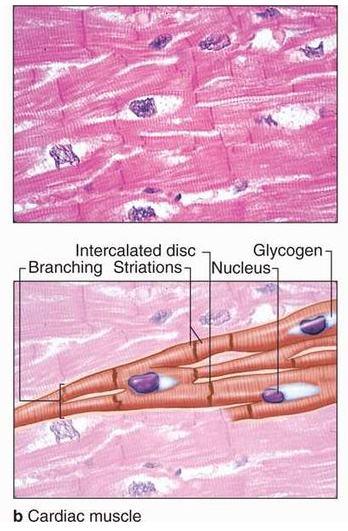
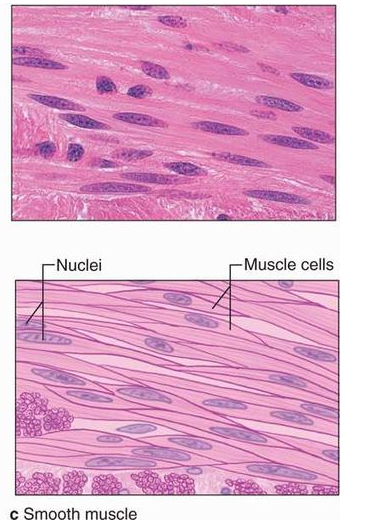
Figure 8: Muscular Tissue
Nervous (Neural) Tissue
Neural Tissue
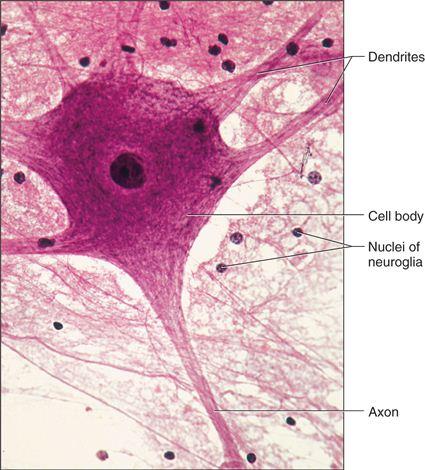
The brain, spinal cord, and nerves all include nerve tissue or neural tissue. It is in charge of coordinating and controlling various bodily functions. It promotes muscle contraction, raises environmental awareness, and is involved in emotions, memory, and thinking. The body's reactivity to changing conditions is mostly controlled by neural tissue. Nerve cells, often known as neurons, make up nervous tissue. Neurons are specialized cells that respond to stimuli by sending out signals via axons, which are elongated projections that emerge from the cell body.
Nervous tissue contains two types of cells namely neurons and neuroglia. Neurons are excitable cells that make up the neural system. A cell membrane surrounds neuronal cells. The nucleus of neurons includes genes. Cytoplasm, mitochondria, and other organelles can be found in neurons. Neurons don't divide in any manner. Nerves are actually neuron projections.The rest of the neural system is made up of neuroglial cells that protect and maintain neurons. Neuroglia accounts for more than half of all neural tissue in our bodies. Neuroglia is non-neuronal cells that do not produce electrical impulses in the central nervous system and peripheral nervous system. Neuroglia are found in both invertebrates and vertebrates' nervous systems and are distinguishable from neurons by the absence of axons and the presence of only one kind of process. Furthermore, they do not establish synapses and keep their potential to divide throughout their lives.When a neuron is activated sufficiently, an electrical disturbance is produced that travels quickly through its plasma membrane. When a disturbance reaches the neuron's ends or output zone, it sets off a chain of events that can stimulate or inhibit neighbouring cells.
The nervous tissue is the main tissue component of the two major parts of the nervous tissue: the central nervous system (CNS), which is formed by the spinal cord and brain, and the peripheral nervous system (PNS), which controls and regulates the body's functions and activities. Nervous tissue can be found in peripheral nerves throughout the body as well as central nervous system components such as the spinal cord and brain.
organ and organ system
Organ and organ system
In multicellular organisms, the fundamental tissues indicated above arrange to form organs, which then associate to form organ systems. Such organization is required for the millions of cells that make up an organism's activities to be more efficient and coordinated. One or more types of tissues make up each organ in our body. Our heart, for example, is made up of all four types of tissues: epithelial, connective, muscular, and neural. After some thorough examination, we also discover that the complexity of organs and organ systems follows a predictable pattern. The term for this discernible pattern is the evolutionary trend. The morphology and anatomy of three species at various evolutionary levels are reviewed below to demonstrate how they are organized and operate.The study of form or externally observable features is known as morphology. The term morphology only refers to this in the case of plants or microorganisms. This refers to the exterior appearance of organs or sections of the body in animals. The term anatomy is commonly used to describe the morphology of internal organs in animals. The earthworm, cockroach, and frog, which represent invertebrates and vertebrates, will be studied for their morphology and anatomy.
Cockroach
Cockroach
Cockroaches are dark or black-bodied animals belonging to the Phylum Arthropoda's class Insecta. Cockroaches with bright yellow, red, and green colorshave also been documented in tropical areas. Their antennae, legs, and a flat extension of the upper body wall that conceals the head vary in size from 14 inches to 3 inches (0.6-7.6 cm). They are nocturnal omnivores who live in moist environments all over the world. They have become occupants of human homes, making them major pests and disease vectors.
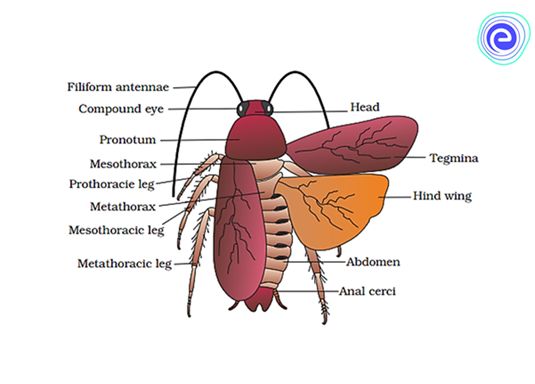
(i)Morphology: The common cockroach, Periplaneta americana, has wings that extend past the tip of the abdomen in males and is around 34-53 mm long inadults. The cockroach's body is divided into three separate regions: the head, the thorax, and the abdomen. A strong chitinous exoskeleton covers the entire body (brown). Hardened plates called sclerites (tergites dorsally and sternites ventrally) are linked to each other by a thin and flexible articular membrane in each segment of the exoskeleton (arthrodial membrane).
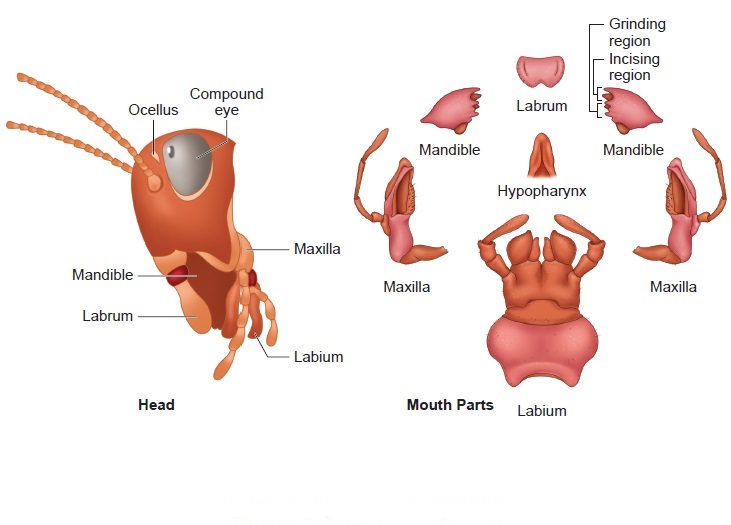
The triangular head is positioned anteriorly at right angles to the longitudinal axis of the body. It is made up of the fusion of six segments and has a flexible neck that allows it to move freely in all directions. A pair of compound eyes can be found on the head capsule. Membranous receptacles in front of the eyes sprout a pair of thread-like antennae. Sensory receptors in antennae aid in environmental monitoring. Appendages on the anterior end of the skull produce biting and chewing mouth parts. A labrum (upper lip), a pair of mandibles, a pair of maxillae, and a labium make up the mouthparts (lower lip). The tongue (hypopharynx) is a flexible lobe that resides within the space encompassed by the mouthparts.
Prothorax, mesothorax, and metathorax are the three sections of the thorax. The neck is a short extension of the prothorax that connects the head to the thorax. Walking legs are attached to each thoracic section. The mesothorax produces the first pair of wings, whereas the metathorax produces the second pair. Tegmina, or opaque dark and leathery forewings (mesothoracic), cover the hind wings when at rest. The rear wings are transparent and membranous, and they help the animal fly.
Both males and females have ten segments in their abdomen. The 7th sternum is boat-shaped in females, and it creates a brood or genital pouch with the 8th and 9th sterna, which contains the female gonopore, spermathecal pores, and collateral glands in the front section.The genital pouch or chamber is located at the back end of the abdomen, between the 9th and 10th terga and the 9th sternum. Dorsal anus, ventral male genital hole, and gonapophysis are all found here. Males have a pair of short, threadlike anal styles that females lack. Anal cerci are a pair of jointed filamentous structures found on the 10th segment of both sexes.
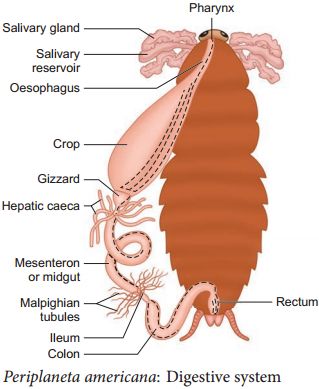
(ii) Anatomy: The alimentary canal is separated into three sections in the body cavity: foregut, midgut, and hindgut. The mouth opens into a short tubular pharynx, which leads to the oesophagus, a tiny tube conduit.
The esophagus expands into a sac-like structure called a crop, which is used to store food. Gizzard or proventriculus follows the crop. It possesses a thick outer layer of circular muscles and a thick inner cuticle that forms six teeth-like chitinous plates. The gizzard assists in the crushing of food particles. The cuticle covers the entire foregut. At the confluence of the foregut and midgut is a ring of 6-8 blind tubules called the hepatic or gastric caeca, which release digestive juice.Another ring of 100-150 yellow-colored thin filamentous Malpighian tubules exists at the midgut-hindgut junction. They aid in excretory product clearance from the hemolymph. The ileum, colon, and rectum make up the hindgut, which is larger than the midgut. Through the anus, the rectum opens up.
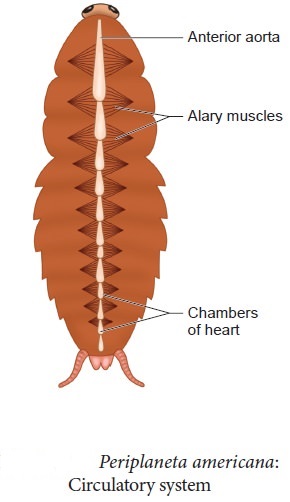
Cockroaches have an open blood circulatory system. The blood vessels are underdeveloped and open into the air (hemocoel). The blood bathes the visceral organs in the haemocoel (hemolymph). Colorless plasma and haemocytes make up the haemolymph. The cockroach's heart is an extended muscular tube that runs along the mid-dorsal line of the thorax and abdomen. It is divided into funnel-shaped chambers on both sides by ostia.Through the ostia, blood from the sinuses enters the heart and is pumped anteriorly to the sinuses. The respiratory system is made up of a network of trachea that open through ten pairs of small openings on the body's lateral side called spiracles. Air is carried to all parts by thin branching tubes (tracheal tubes separated into tracheoles). The sphincters control the opening of the spiracles.
Diffusion occurs at the tracheoles to exchange gases. Malpighian tubules are responsible for excretion. Glandular and ciliated cells line each tubule. They take in nitrogenous waste and convert it to uric acid, which is then expelled through the hindgut. As a result, It is known as uricotelic.
The fat body, nephrocytes, and urecose glands all contribute to excretion. On the ventral side, the cockroach's nervous system is made up of a series of fused, segmentally arranged ganglia connected by paired longitudinal connectives. The thorax has three ganglia and the abdomen has six. Cockroaches have a neurological system that runs throughout their bodies. A portion of the nervous system is located in the skull, while the rest is located on the ventral (belly) side of the animal. Because of this, even if a cockroach's head is removed, it can survive for up to one week. The brain is represented in the head area by the supra-oesophageal ganglion, which sends nerves to the antennae and compound eyes.
Antennae, eyes, maxillary palps, labial palps, anal cerci, and other sensory organs are found in cockroaches. The complex eyes are located on the head's dorsal surface. About 2000 hexagonal ommatidia make up each eye. A cockroach can obtain several photos of an object by using many ommatidia. Mosaic vision is a type of vision that has a higher sensitivity but lower resolution and is common at night (hence called nocturnal vision).
Cockroaches are dioecious, with well-developed reproductive organs in both sexes. A pair of testes, one on each lateral side in the 4th -6th abdominal segments, make up the male reproductive system.
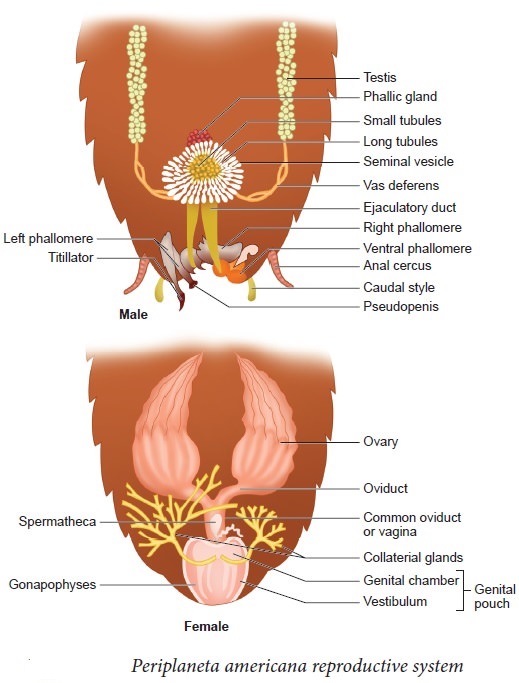
A narrow vas deferens develops from each testis and opens into the ejaculatory duct via the seminal vesicle.The male gonopore is located ventral to the anus and opens into the ejaculatory duct. In the 6th and 7th abdominal segments, there is a mushroom-shaped gland that serves as an accessory reproductive gland. Male gonapophysis or phallomeres depict the external genitalia (chitinous asymmetrical structures, surrounding the male gonopore). The sperm are retained in the seminal vesicles and cemented together in spermatophores, which are expelled during copulation. Two enormous ovaries lie laterally in the 2nd – 6th abdominal segments, forming the female reproductive system. Each ovary is made up of eight ovarian tubules, also known as ovarioles, which contain a chain of growing ova. Each ovary's oviducts join to form a single median oviduct (also known as the vagina), which exits into the genital chamber.The 6th segment, which enters the vaginal chamber, contains a pair of spermatheca. Spermatophores are used to transport sperm. The fertilized eggs are enclosed in oothecae capsules. The ootheca is a 3/8" (8 mm) length dark reddish to a blackish-brown capsule. They're dropped or bonded to a suitable surface, usually near a food source in a crack or crevice with high relative humidity. Females produce 9-10 oothecae on average, each holding 14-16 eggs. P. americana has a paurometabolous development, which means it develops through the nymphal stage. The nymphs have the appearance of adults. To attain adulthood, the nymph must molt 13 times.Wing pads are present in the next to last nymphal stage, however, only adult cockroaches have wings.
Many cockroach species are found in the wild and have yet to be identified as economically important. In and around the human habitat, a few species flourish. They are pests because their stinky excreta spoils food and contaminates it. By polluting food, they can spread a range of bacterial infections.
earthworm
Earthworm
The earthworm is a reddish-brown terrestrial invertebrate that lives in the moist soil's upper layer. They spend the day in tunnels created by burrowing and swallowing earth. Worm castings, or faecal deposits, are used to track them down in the gardens. Pheretima and Lumbricus are the most prevalent Indian earthworms.
(i) Morphology: Earthworms' bodies are long and cylindrical. The body is divided into over a hundred short, identical pieces (metameres about 100-120 in number). Along the longitudinal axis of the body, a dark median mid dorsal line (dorsal blood vessel) marks the dorsal surface. The presence of genital apertures distinguishes the ventral surface (pores).
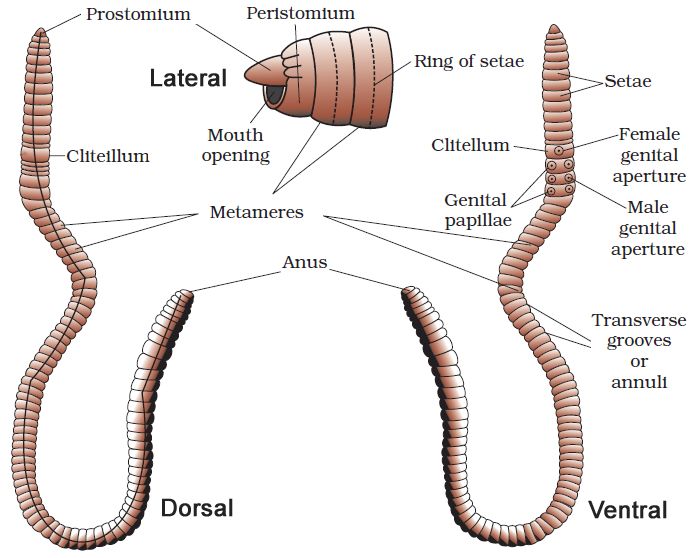
The mouth and the prostomium, a lobe that serves as a mouth cover and a wedge to force open soil crevices into which the earthworm may crawl, make up the anterior end. The role of the prostomium is sensory. The peristomium (buccal segment) is the first body segment and houses the mouth. A conspicuous dark band of glandular tissue termed clitellum covers segments 14-16 in a mature worm. As a result, the body is divided into three distinct regions: preclitellar, clitellar, and postclitellar. The ventrolateral sides of the intersegmental grooves, i.e. the 5th -9th segments, have four pairs of spermathecal openings. In the mid-ventral line of the 14th segment, there is a solitary female genital pore.On the ventrolateral sides of the 18th segment, there are two male genital pores. Nephridiopores are tiny pores that open on the surface of the body. Except for the first, last, and clitellum, each body segment has rows of S-shaped setae inserted in the epidermal pits in the middle. Setae can be retracted or expanded. Their primary function is movement.
(ii) Anatomy: A thin non-cellular cuticle covers the earthworm's body wall, which is followed by the epidermis, two muscle layers (circular and longitudinal), and an innermost coelomic epithelium. The epidermis is a single layer of columnar epithelial cells with secretory gland cells.
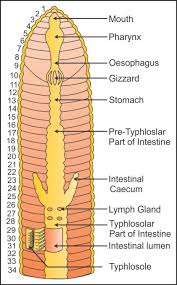
The alimentary canal is a straight tube that connects the first and last body segments. The buccal cavity (1-3 segments) leads into the muscular pharynx through a terminal mouth. The esophagus (5-7 segments) is a tiny, narrow tube that leads to the muscular gizzard (8-9 segments). It aids in the grinding of soil particles and rotting leaves, among other things. The stomach is divided into 9-14 parts. Earthworms eat decaying leaves and organic debris mixed with soil as nourishment.The humic acid in the humus is neutralized by calciferous glands in the stomach. The stomach begins at the 15th section and continues until the last part. On the 26th segment, a pair of short and conical intestinal caecae protrude from the gut. Except for the last 23rd-25th segments, the existence of an internal median fold of the dorsal wall called typhlosole is a distinguishing feature of the intestine after the 26th segment. This expands the absorption surface area in the gut. A little spherical opening called the anus connects the alimentary canal to the outside world. Digestive enzymes break down complicated meals into tiny absorbable components when the organic-rich soil moves through the digestive tract. These simpler molecules are absorbed and used through intestinal walls.
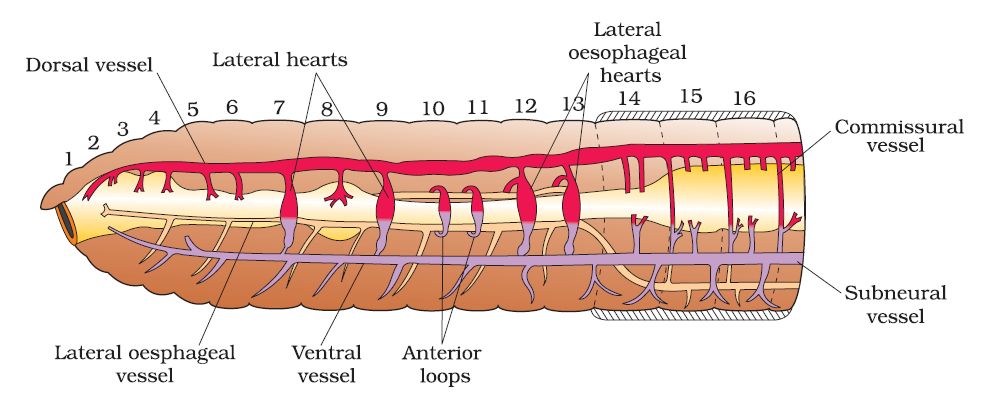
The blood vascular system of Pheretima is closed, consisting of blood arteries, capillaries, and the heart. Blood is restricted to the heart and blood vessels due to the closed circulatory system. Blood circulates in one direction due to contractions. The gut, nerve cord, and body wall are all supplied by smaller blood vessels. The 4th, 5th, and 6th segments all have blood glands. They manufacture blood cells as well as haemoglobin, which is dissolved in blood plasma. Blood cells are naturally phagocytic.
Earthworms do not have specific breathing apparatus. The wet body surface allows for respiratory exchange into the bloodstream.
Nephridia are segmentally arranged coiled tubules that make up the excretory organs (sing.: nephridium). There are three types of nephridia:
(i) Septal nephridia, which are present on both sides of intersegmental septa from segment 15 to the last that open into the intestine,
(ii) Integumentary nephridia, which are attached to the lining of the body wall from segment 3 to the last that open on the body surface, and
(iii) Pharyngeal nephridia, present as a three-paired outfit in 4th, 5th and 6th segment. The structure of these many nephridia is remarkably similar. The volume and composition of the bodily fluids are regulated by nephridia. A nephridium begins as a funnel, collecting surplus fluid from the coelomic chamber.The funnel connects to a tubular component of the nephridium, which transports wastes to the digestive tube through a pore in the body wall.
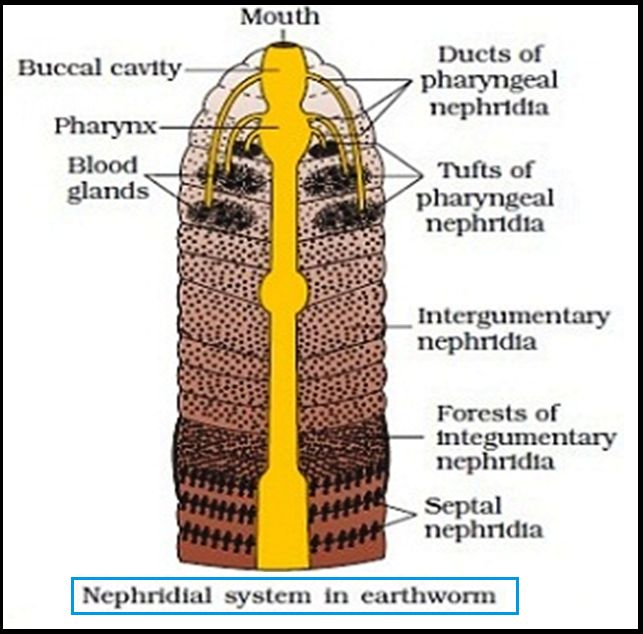
Ganglia are positioned segment-wise on the ventral paired nerve cord to represent the nervous system. In the front region (3rd and 4th segments), the nerve cord bifurcates, laterally enclosing the pharynx and dorsally joining the brain ganglia to form a nerve ring. The cerebral ganglia, along with other nerves in the ring, integrate sensory input and control bodily muscle responses.Although the sensory system lacks eyes, it does have light and touch-sensitive organs (receptor cells) that allow it to discern between light intensities and feel ground vibrations. Chemoreceptors (taste receptors) in worms are specialized to respond to chemical inputs. These sensory organs are found on the worm's front end.
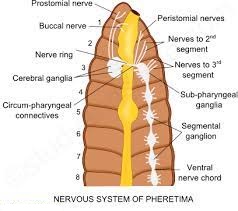
Earthworms are hermaphrodites (bisexual), which means they have both testes and ovaries. In the 10th and 11th segments, there are two pairs of testes. Their vasa deferentia connect to the prostatic duct at the 18th segment. In the 17th and 19th segments, there are two pairs of auxiliary glands, one pair in each.A pair of male genital holes on the ventrolateral side of the 18th segment access the outside the shared prostate and spermatic duct (vasa deferentia). In the 6th-9th segments, there are four pairs of spermathecae (one pair in each segment). During copulation, they receive and store spermatozoa. At the inter-segmental septum of the 12th and 13th segments, one pair of ovaries is attached. Underneath the ovaries are ovarian funnels that continue into the oviduct, unite, and open on the ventral side as a single median female genital hole on the 14th segment.
During the mating process, two worms exchange sperm. One worm must discover another worm, and they mate by exchanging spermatophores (packets of sperm) at opposite gonadal apertures.Cocoons made by clitellum gland cells contain mature sperm and egg cells, as well as nourishing fluid. Fertilization and development take place within the soil-deposited cocoons. The sperm cells within the cocoon fertilize the ova (eggs), which subsequently fall off the worm
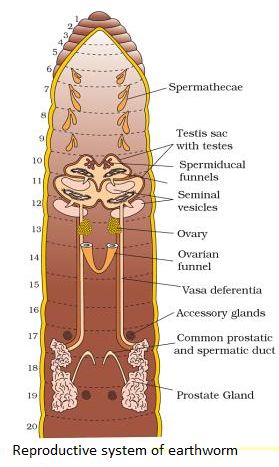
and land in or on the earth. The worm embryos are kept in the cocoon. Each cocoon generates two to twenty baby worms on average after about three weeks. Earthworms develop directly, without the formation of a larva. Earthworms are known as "farmer's friends" because they dig burrows in the soil and make it porous, which aids in plant root penetration and respiration.Vermicomposting is the method of using earthworms to increase soil fertility. They're also utilized as game fishing bait.
frog
Frog
Frogs are members of the phylum Chordata's class Amphibia, which can live on land and in freshwater. Rana tigrina is the most common frog species in India. They do not have a steady body temperature; rather, their body temperature changes depending on the ambient temperature. Cold-blooded animals are known as poikilotherms. You may have also noticed that the frogs' color changes while they are in grasses or on dry soil. They can change their color to hide from their adversaries (camouflage). The term for this protective coloring is mimicry. During the summer and winter, the frogs are not visible. They take refuge in underground tunnels during this time to avoid the excessive heat and cold.Summer sleep (aestivation) and winter sleep (hibernation) are the two types of sleep.
(i) Morphology: Due to the presence of mucus, the skin is smooth and slippery. Moisture is always retained on the skin. The dorsal side of the body is olive green in color with dark irregular markings. The skin is uniformly pale yellow on the ventral side. The frog does not drink water; instead, it absorbs it through its skin. A frog's body is divided into two parts: head and trunk. There is no neck or tail. A pair of nostrils can be found above the mouth. The eyes protrude and are protected by a nictitating membrane while in the water. A membranous tympanum (ear) receives sound signals on either side of the eyes. Swimming, walking, leaping, and digging are all assisted by the forelimbs and hindlimbs.The hind limbs have five fingers and are larger and more muscular than the forelimbs, which have four. Webbed digits on the feet aid with swimming. Sexual dimorphism is seen in frogs. Male frogs can be recognized from female frogs by the presence of sound-producing vocal sacs and a copulatory pad on the first digit of the forelimbs.

(ii) Anatomy: Frogs' bodily cavities house many organ systems with well-developed structures and functions, including the digestive, circulatory, respiratory, nervous, excretory, and reproductive systems. The alimentary canal and digestive glands make up the digestive system. Because frogs are carnivores, the alimentary canal is short, resulting in a shorter gut. The pharynx connects the mouth to the buccal cavity, which leads to the esophagus. The esophagus is a short tube that connects the mouth to the stomach, then to the intestine, rectum, and finally to the cloaca. Bile is produced by the liver and stored in the gall bladder. Pancreatic juice, which contains digestive enzymes, is produced by the pancreas, a digestive gland. The bilobed tongue is responsible for capturing food.
Food is broken down by the action of Hydrochloric acid and gastric secretions released from the stomach's walls. Chyme, or partially digested food, is transferred from the stomach to the duodenum, the first segment of the small intestine. Through a shared bile duct, the duodenum receives bile from the gall bladder and pancreatic secretions from the pancreas. Pancreatic fluids digest carbs and proteins while bile emulsifies fat. The intestine is where the final digesting takes place. The many finger-like folds in the inner wall of the intestine called villi and microvilli absorb digested food. The undigested solid waste goes through the cloaca and into the rectum. Frogs have two different strategies to breathe on land and in water.
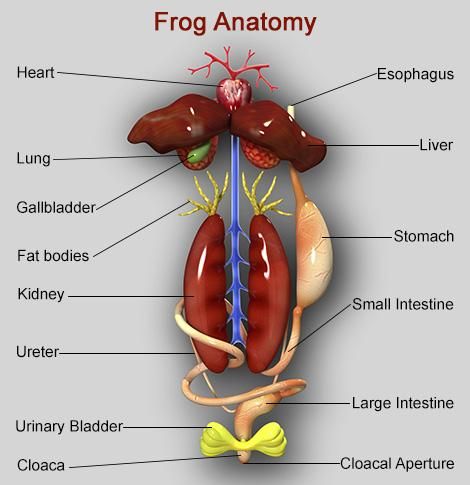
Skin serves as an aquatic respiratory organ in water (cutaneous respiration). Diffusion exchanges dissolved oxygen in the water through the skin. The buccal cavity, skin, and lungs serve as respiratory organs on land. Pulmonary respiration is the process of breathing through the lungs. The lungs are a pair of elongated, pink-colored sac-like structures located near the top of the trunk (thorax). Air enters the lungs via the nostrils, buccal cavity, and lungs. Gaseous exchange occurs through the skin during aestivation and hibernation. Frogs have a well-developed closed circulatory system. Frogs, like humans, have a lymphatic system. The heart, blood arteries, and blood make up the blood vascular system. Lymph, lymph channels, and lymph nodes make up the lymphatic system.
The heart is a muscle structure located in the upper body cavity. It has three chambers, two atria, and one ventricle, and is surrounded by a pericardium membrane. The right atrium is connected to the sinus venosus, a triangle structure. It receives blood through the vena cava, which are large veins. On the ventral side of the heart, the ventricle opens into a sac-like conus arteriosus. The arteries transport blood from the heart to all regions of the body (arterial system). The venous system collects blood from various regions of the body and transports it to the heart.
Frogs have a unique venous connection between the liver and gut, as well as the kidney and lower body.The first is known as the hepatic portal system, whereas the second is known as the renal portal system. Plasma and cells are found in the blood. RBCs (red blood cells) or erythrocytes, WBCs (white blood cells) or leucocytes, and platelets are the blood cells. RBCs are nucleated and contain the pigment haemoglobin, which is red in color. Lymph is not the same as blood. It is deficient in proteins and RBCs. During circulation, the blood transports nutrients, gases, and water to their proper locations. The pumping activity of the muscular heart is responsible for blood circulation. A well-developed excretory system is responsible for the removal of nitrogenous wastes. A pair of kidneys, ureters, cloaca, and urine bladder make up the excretory system.
On both sides of the vertebral column, they are compact, dark red, bean-like structures located a bit posteriorly in the body cavity. Uriniferous tubules or nephrons are structural and functional units that make up each kidney. Male frogs have two ureters that arise from their kidneys. The ureters serve as a urinogenital channel, leading to the cloaca. The ureters and oviduct open separately in the cloaca in females. The rectum, which likewise opens in the cloaca, has a thin-walled urine bladder ventral to it. The frog is a ureotelic animal because it excretes urea. Blood transports excretory wastes to the kidney, where they are sorted and expelled. The frog's control and coordination system is highly developed.
It involves both the nervous and endocrine systems. Hormones released by the endocrine glands are responsible for the chemical coordination of the body's many organs. The pituitary, thyroid, parathyroid, thymus, pineal body, pancreatic islets, adrenals, and gonads are the most significant endocrine glands in frogs. The nervous system is arranged into a central nervous system (brain and spinal cord), a peripheral nervous system (cranial and spinal nerves), and an autonomic nervous system (sympathetic and parasympathetic) (sympathetic and parasympathetic). The brain gives rise to ten pairs of cranial nerves. The brain is protected by a bony structure known as the brain box (cranium). The forebrain, midbrain, and hindbrain are the three parts of the brain. The olfactory lobes, paired cerebral hemispheres, and unpaired diencephalon makes up the forebrain.
A pair of optic lobes distinguish the midbrain. The cerebellum and medulla oblongata make up the hindbrain. The medulla oblongata exits the foramen magnum and continues into the spinal cord, which is contained within the vertebral column. The sense organs of the frog are touch (sensory papillae), taste (taste buds), smell (nasal epithelium), vision (eyes), and hearing (ears) (tympanum with internal ears). Eyes and internal ears are well-organized structures, with the rest consisting of cellular aggregations around nerve endings. Frog eyes are a pair of spherical structures in the orbit of the skull. These are basic eyeballs (possessing only one unit). In frogs, the external ear is missing, leaving just the tympanum visible.
The ear is both a hearing and a balancing organ (equilibrium). Male and female reproductive systems in frogs are well-organized. A pair of yellowish ovoid testes are discovered adhering to the top section of the kidneys by a double fold of peritoneum called the mesorchium in male reproductive organs. The number of Vasa efferentia that emerge from the testes is 10-12. On their side, they enter the kidneys and open into Bidder's canal. Finally, it connects with the urinogenital duct, which runs from the kidneys to the cloaca. The cloaca is a small, midline chamber where faeces, urine, and sperms are passed to the outside. A pair of ovaries are among the female reproductive organs.
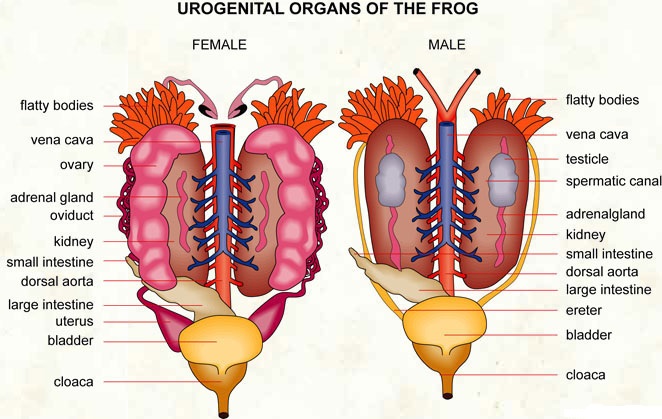
The ovaries are close to the kidneys but have no functional relationship with them. Separate oviducts emerge from the ovaries and open into the cloaca. At any given time, a mature female can lay between 2500 and 3000 ova. External fertilization takes place in water. Tadpoles are in the larval stage of development. Tadpoles go through metamorphosis to become adults. Frogs are advantageous to humans because they consume insects and protect crops. Frogs keep the ecology in balance since they are a vital link in the food chain and food web. Humansconsume the muscular legs of frogs in several regions.
