 ACME SMART PUBLICATION
ACME SMART PUBLICATION
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
TRANSPIRATION
Transpiration is the loss of water in the form of vapours from the aerial parts of the plant.
The loss of water is no great that it reduces water level in the soil and can lead to e ea of plant, but transpiration is said to be necessary for water and mineral absorption, ascent of sap and lowering the temperature (cooling effect).
So, transpiration is called as necessary evil (Curtis) or an unavoidable evil (Steward).
About 98% of water absorbed by land plants evaporate from their aerial parts and is lost into the atmosphere.
Less than 1 % of the water reaching the leaves is used in photosynthesis and plant growth.
Types of Transpiration
Based on the plant parts or structure involved, following four types can be recognised:
1. Stomatal transpiration: It is the transpiration that occurs through the stomata. The epidermis of leaves and green stems have numerous stomata. These are responsible for about 50 -97% of the total water transpired.
2. Cuticular transpiration : Water vapours are also lost directly from the outer walls of the epidermal cells through the cuticle. Cuticle is a wax like layer of cutin that covers the epidermis of leaves and stems. It reduces the water loss but may give out water vapours through the cracks. It commonly constitutes 3-10% of total transpiration. It is maximum upto 50% in herbaceous plants, ferns etc. growing in shady places.
3. Lenticular transpiration : Lenticels are aerating pores in the cork of the woody stems, twigs and fruits. Water vapours are lost through these openings. The amount of water vapours lost through lenticels is usually insignificant (approximately 0.1% of the total water loss).
4. Bark transpiration: This occurs through the bark of woody stem. It contributes about 1% of the total transpiration.
Structure of Stomata
Stomata are the tiny apertures found on the epidermis of leaves and young green stems.
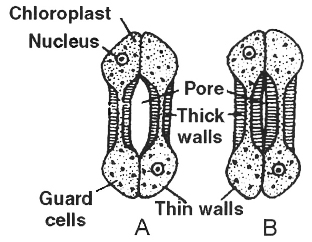
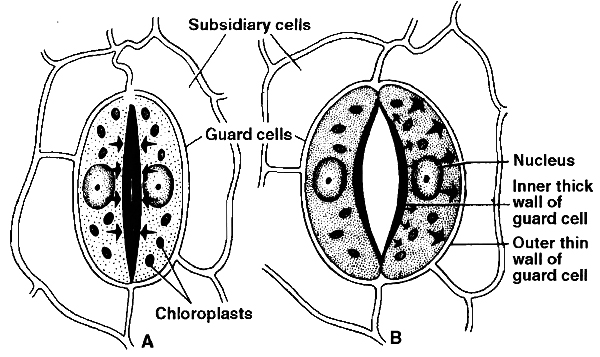
Each stoma is surrounded by two specialized epidermal cells, called guard cells.
They differ from epidermal cells in their shape (kidney or bean shaped) and in the presence of chloroplasts.
The inner wall of the guard cell is thick and elastic, whereas the outer wall is thin.
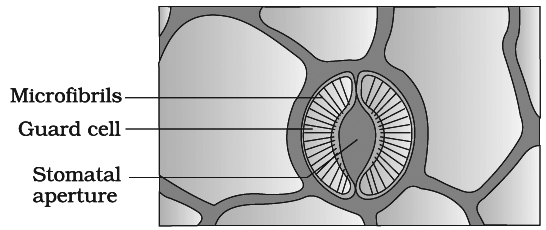
Opening of the stoma is also aided due to the orientation of the microfibrils in the cell walls of the guard cells.
Cellulose microfibrils are oriented radially rather than longitudinally, making it easier for the stoma to open.
Guard cells are bordered by one or more modified epidermal cells called subsidiary cells or accessory cells.
In monocots, guard cells are ellipsoidal or dumb-bell shaped (called graminaeous stomata or poaceous stomata).
These stomata have thin end walls and thick walled middle region.
The leaf surface, depending upon the species may contain 1000 to 60,000 stomata per square centimeter.
The total pore area is approximately 1 -2% of total leaf area.
Classification of stomata on the basis of their distribution
With the exception of few submerged hydrophytes, stomata are widely distributed amongst angiosperms and gymnosperms.
Based on their distribution on leaf surface, stomata are grouped in the following five categories:
1. Apple or mulberry type: Stomata are present only on the lower leaf surface, e.g., apple. Such leaves are called hypostomatic.
2. Potato type : The stomata occur on both the leaf surfaces, being more on the lower surface than that on the upper, e.g., potato, tomato, brinjal. Such leaves are called amphistomatic.
3. Oat type: The stomata occur equally on both the leaf surfaces, e.g., wheat, rice, grasses, etc. These leaves are also called amphistomatic.
4. Water lily type: The stomata are found only on the upper surface of the leaf, e.g., water lily. The leaves of such plants are found floating on water surface. These leaves are termed as epistomatic.
5. Potamogeton type: The stomata in some hydrophytes are either absent or vestigeal. Such leaves are called astomatic.
Concept Builder
Classification of Stomata on the basis of daily movement
Loftfield classified stomata into following four types on the basis of their daily movement:
(a) Alfalfa type. The stomata remain open throughout the day and closed throughout the night, e.g., pea, beans, radish, mustard, turnip, grapes etc.
(b) Potato type. The stomata remains open throughout the day and night except for a few hours in the evening e.g., onion, potatoes, cabbage, banana, etc.
(c) Barley type. The stomata open for only a few hours during the day and remain closed for rest of the period, e.g., barley, maize, wheat and other cereals.
(d) Equisetum type. In Equisetum (Horsetail), the stomata remain open throughout the day and night
Mechanism of Opening and Closing of Stomata
Stomata function as turgor operated valves.
When osmotic concentration of guard cells increases, water comes in and guard cells become turgid and stomata gets open.
Whenever, osmotic concentration of guard cells decreases water moves out, guard cells become flaccid and hence get closed.
This increase and decrease in osmotic concentration is explained by a number of theories, described below:
1. Photosynthetic Theory (Von Mohl and Schwendener) :
This theory proposes that in the morning, as soon as light is available chloroplasts of guard cell start photosynthesis, as a result sugars are produced, which increases the osmotic concentration of guard cells, water comes in, guard cells become turgid and stomata are open.
This theory is not accepted because photosynthetic activity of guard cell chloroplasts seems to be negligible and sugar does not occur in detectable quantity in guard cells (as Rubisco is absent in guard cell chloroplast).
2. Starch ![]() Sugar hypothesis (Classical theory).
Sugar hypothesis (Classical theory).
It was given by Sayre and Scarth and later modified by Steward.
This theory is called classical theory.
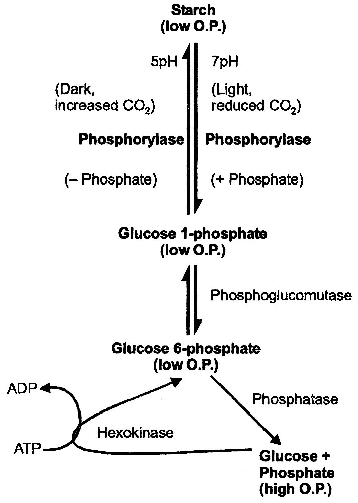
They indicated that interconversion of starch into sugar is determined by change in pH.
According to this theory, carbon dioxide liberated due to respiration is used in photosynthesis by mesophyll cells.
This results in increase in pH to 7-7.5.
In this alkaline state, starch is converted to glucose-1-phosphate.
In the dark, carbon dioxide accumulates in the intercellular spaces as it is not utilised in photosynthesis.
It lowers pH of the guard cells to about 5.
In this acidic state, glucose-1-phosophate is converted back to starch, leading to closure of stomata.
in guard cells according to Starch Sugar Hypothesis
Objectivess
(i) Starch is absent in onion.
(ii) Glucose does not appear in detectable quantity in guard cells.
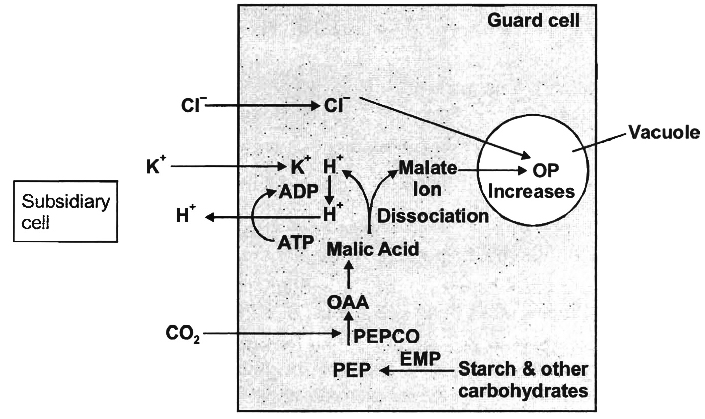
3. Active K+ transport or Potassium pump theory.
Two Japanese scientists, S. Imamura and M. Fujino showed the accumulation of K+ in the guard cells during stomatal opening.
Later, Levitt (1974) explained the influx of K+ ions in the guard cells and their critical role in stomatal movement.
(a) Opening of stomata in light:
(i) In light, starch in the guard cells is incompletely oxidized into phosphoenol pyruvate (PEP). It is later converted into or anic acids, particularly malic acid. This reaction is catalyzed by an enzyme phosphoenol pyruvate carboxylase (PEPCO or PEPCase).
(ii) Malic acid dissociates into malate ion and protons (H+) in the guard cells.
(iii) H+ from guard cells, are transported to epidermal cells and K+ from epidermal cells gets into the guard cells through the agency of hydrogen-potassium ion exchange pump in the plasma membrane.
(iv) In the guard cells, K+ ions are balanced by malate anions. Besides, small amount of Cl– ions are also absorbed which neutralize a small percentage of K+ ions.
(v) The process of ion exchange requires ATP and thus, it is an active process.
(vi) Increased K+ and malate ions forms potassium malate and store it in vacuoles of the guard cells, increasing their osmotic concentration. Hence, water enters the guard cells by endosmosis.
(vii) Turgor pressure of the guard cells increases due to endosmosis and the stomata gets open.
(b) Closing of stomata in the dark:
(i) As CO2 is not utilized in photosynthesis during night, hence its concentration in the sub stomatal cavity increases.
(ii) An inhibitor hormone-abscissic acid (ABA) functions in the presence of CO2, It inhibits K+ ion uptake by changing the diffusion and permeability of the guard cells for positive ions.
(iii) The K+ ions are transported back to the epidermal or subsidiary cells from the guard cells.
Concept Builder
In succulents stomata open during the night and close during the day. This is called scotoactive opening. These plants show night time formation of malic acid, e.g., Opuntia, Bryophyllum.
Factors affecting Transpiration
(A) External factors
(1) Light. Blue light induces maximum opening of stomata. In its absence, stomata remain closed. Light also affects the rate of transpiration by increasing temperature. Blue and red light are effective for transpiration, constituting its action spectrum.
(2) Relative humidity (vapour pressure gradient). In humid atmosphere (when the relative humidity is high) the rate of transpiration decreases. In dry atmosphere the relative humidity is low (low water vapour pressure), so the rate of transpiration increases.
(3) Temperature. Higher the temperature more is the rate of transpiration as it results in a higher vapour pressure gradient. Lowering of temperature decreases the rate of transpiration.
(4) Wind. If wind is not blowing, water vapours accumulate above the transpiring leaves which decrease the rate of transpiration. The blowing wind removes the accumulated humidity and brings fresh air capable of absorbing water (slow breeze) and thus, the rate of transpiration is enhanced.
Storm or wind of very high velocity causes closure of stomata, thus checking transpiration.
(5) Available soil water. If the available water in the soil is not sufficient, the rate of transpiration is decreased. A high concentration of salts in the soil water also reduces the rate of transpiration due to less water absorption.
(B) Plant Factors
1. Root-shoot ratio: Root absorbs water, hence if root shoot ratio decreases then transpiration decreases or vice versa. Short plants have a high root -shoot ratio.
2. Structure of leaf: Many features like thick cuticle, waxy coating, thick walled hypodermis, sunken stomata reduce the transpiration.
3. Number and distribution of open stomata, plant water, canopy structure are other factors affecting transpiration.
Significance of Transpiration
Transpiration has been described as a 'necessary evil' (Curtis) and an unavoidable evil (Steward). It is potentially harmful.
The stomata remain open for exchange of gases.
This also results in the loss of water as vapours.
Consequently, the water level in the soil is reduced, causing wilting of plants.
The stomata, however, cannot be closed to prevent water loss because this would also stop gaseous exchange needed for respiration and photosynthesis.
The advantages of transpiration are as follows:
(i) The absorption of water and ascent of sap to various parts of the plant body is mostly due to transpiration. Transpiration pull is responsible for mass movement of water and solutes in upward direction
(ii) Plants receive solar energy in very large amounts for the synthesis of carbohydrates. If there is no dissipation of energy, the temperature of leaf surface would rise to a lethal level in less than two minutes. Transpiration plays an important role here. It helps in dissipating this excess energy by evaporating water from the leaf surface and thus, helps in keeping the plant cool.
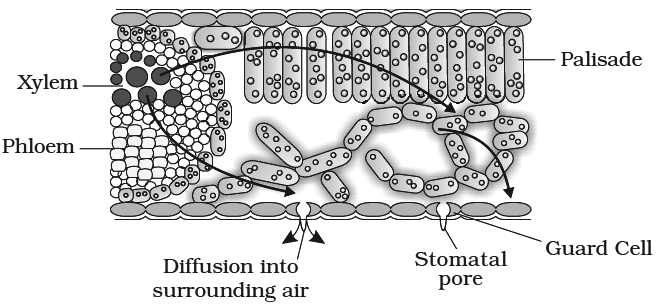
between the outside air and the air spaces of the leaf. The gradient is transmitted into the photosynthetic cells and on the water-filled xylem in the leaf vein.
(iii) Development of mechanical tissues, growth of root system, increasing ash and sugar content of fruits and development of resistance are other beneficial effects of transpiration.
Many chemicals (antitranspirants) have been found to reduce the rate of transpiration without affecting CO2 uptake e.g., Phenyl mercuric acetate (PMA, a fungicide), Abscisic acid (ABA) and CO2, Silicon emulsion and low viscosity waxes cover stomata as a film, allow CO2 and O2 exchange but resist diffusion of water.
Transpiration and Photosynthesis — a Compromis
Transpiration has more than one purpose; it
Creates transpiration pull for absorption and transport in plants.
Supplies water for photosynthesis.
Transports minerals from the soil to all parts of the plant
Cools leaf surfaces, sometimes 10 to 15 degrees, by evaporative cooling.
Maintains the shape and structure of the plants by keeping cells turgid.
An actively photosynthesising plant has an insatiable need for water.
Photosynthesis is limited by available water which can be swiftly depleted by transpiration.
The humidity of rainforests is largely due to this vast cycling of water from root to leaf to atmosphere and back to the soil.
The evolution of the C4 photosynthetic system is probably one of the strategies for maximising the availability of CO2 while minimising water loss.
C4 plants are twice as efficient as C3 plants in terms of fixing carbon (making sugar).
However, a C4 plant loses only half as much water as a C3 plant for the same amount of CO2 fixed.
