ACME SMART PUBLICATION
ACME SMART PUBLICATION
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
THE SEED
Morphologically, ripened ovule after fertilisation is known as seed. Seeds are characteristic of spermatophytes (Gymnosperms and Angiosperms).
Parts of Seed
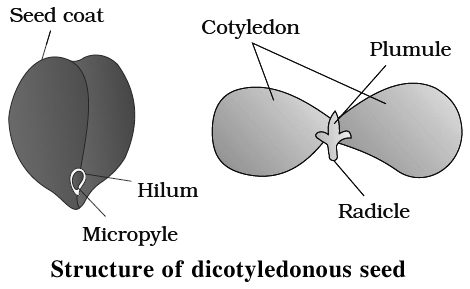
1. Seed coat
Outer, protective covering of the seed is called seed coat, which develops from integuments of ovule.
In seeds developing from bitegmic ovules, there are two distinct layers in seed coat.
The outer layer is thick, hard and leathery (developing from outer integument), called testa, whereas inner layer is thin and papery (developing from inner integument), called tegmen.
In seeds, developing from unitegmic ovules there is single seed coat.
All the structures inside seed coat constitute kernel, while Hilum is a scar on seed coat through which the developing seeds are attached to the fruit.
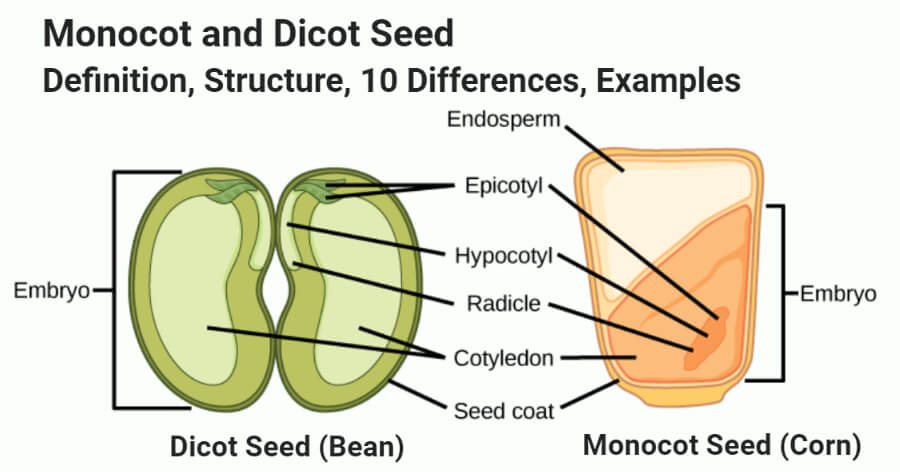
2. Embryo
Embryo is the most important part of the seed, which represents tiny future plant.
The embryo is having an embryonal axis or main axis called tigellum, to which one or two cotyledons (seed leaves) are attached, depending upon whether the seed is monocot or dicot.
The portion of embryonal axis or tigellum below the point of attachment of cotyledons, is called hypocotyl, which bears radicle or future root at its tip.
Similarly, portion of embryonal axis or tigellum above the point of attachment of cotyledons, is called epicotyl, which bears plumule (future shoot) at its tip.
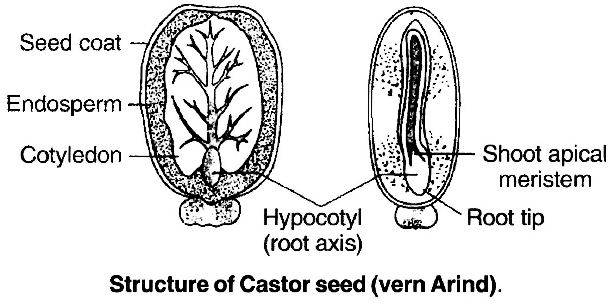
In castor seed (Ricinus communis), there is a specific outgrowth called caruncle present over hilum. It is formed by proliferation of cells of outer integument at tip. Caruncle is somewhat spongy and helps in absorption of water during germination of seed.
Based upon presence or absence of endosperm the seed may be of two types:
(1) Non-endospermic or exalbuminous seeds: In seeds like gram, pea, groundnut, the endosperm is completely consumed by the embryo, thus the seeds are called non-endospermic or exalbuminous e.g., dicots.
(2) Endospermic or albuminous seed: In monocots and castor bean (dicots) embryo does not consume all endosperm. So it persists in the mature seed. Such seeds are called endospermic or albuminous seeds. In these seeds food is stored in endosperm.
Perispermic seeds:
Mostly nucellus is consumed after fertilization due to absorption of food by the endosperm and embryo.
Sometimes, the nucellus remains persistent in the seed and is called perisperm.
Such seeds are called perispermic seeds, e.g., Piper nigrum (black pepper).
Chalazosperm is perisperm like tissue in chalazal region. It is a substitute for endosperm e.g., Cynastrum.
Internal structure of maize seed
On the outside of the grain is present a single thin but hard covering.
It is formed by the fusion of the seed coat or testa and the fruit wall or pericarp.
Below the grain covering are present two structures, endosperm and embryo.
The endosperm consists of two parts, horny aleurone and mealy storage.
The aleurone region lies immediately below the grain covering.
The cells have thick walls and dense cytoplasm filled with aleurone or protein grains.
The latter produce enzymes during the process of grain germination.
The storage region of endosperm is whitish or yellowish.
It is rich in starch grains.
The embryo occurs in the pointed part of the grain, mostly towards the upper side.
It consists of an embryo axis containing a radicle, a plumule and a single, shield shaped, lateral cotyledon.
It has two protective sheaths, radicle sheath is called coleorhiza and a conical protective sheath of plumule is known as coleoptile.
Coleoptile has a terminal pore for the emergence of first leaf during germination.
The sheath is capable of growth.
It assists the future shoot in passing through the soil during germination.
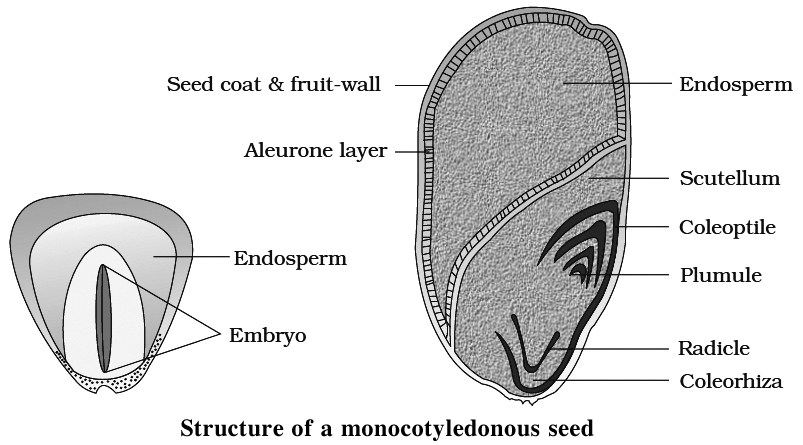
The single cotyledon of Maize grain is called scutellum.
It occupies the major portion of the embryo region of the grain.
The outermost layer at the boundary of endosperm and embryo is known as epithelial layer.
It is both secretory and absorptive.
The epithelial layer secretes hormones into the endosperm for the synthesis of enzymes required for solubilisation of food.
The solubilised food is absorbed by it and then transferred to the embryo axis.
On opposite side of scutellum is present a tongue shaped flap like outgrowth called epiblast, it represent the remains of second cotyledon.
The Seed
After fertilization, the ovules mature into seeds. A seed consists of two parts: a seed coat and an embryo. A radicle, an embryonal axis, and one (as in wheat, maize) or two cotyledons make up the embryo (as in gram and pea).
Structure of a Dicotyledonous Seed:
The seed coat is a seed's outermost coating. The outer testa and the inner tegmen are the two layers of the seed coat. The hilum is a scar on the seed coat that was used to adhere the developing seeds to the fruit.The micropyle is a tiny pore located above the hilum. The embryo, which consists of an embryonal axis and two cotyledons, is contained within the seed coat. The cotyledons are usually plump and rich in food reserves. The radicle and the plumule are located at the two ends of the embryonal axis. The endosperm developed as a result of multiple fertilization in some seeds, such as castor, is a food-storing tissue and is referred to as endospermic seeds.The endosperm is absent in mature seeds of plants such as beans, gram, and pea, and these seeds are referred to as non-endospermic.
Structure of Monocotyledonous Seed:
Monocotyledonous seeds are usually endospermic, but some, such as those found in orchids, are not. The seed coat of cereal seeds, such as maize, is membranous and usually united with the fruit wall. The endosperm is a thick substance that stores food. A protein layer termed aleurone separates the embryo from the endosperm's outer covering. The embryo is tiny and rests in a groove on one end of the endosperm. It consists of a scutellum, a big shield-shaped cotyledon, and a short axis with a plumule and radicle. Coleoptile and coleorhiza are sheaths that surround the plumule and radicle, respectively.