ACME SMART PUBLICATION
ACME SMART PUBLICATION
The Tissues
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
THE TISSUES
The term tissue was coined by Grew.
A group of similar or dissimilar cells that perform a common function and have a common origin is called tissue.
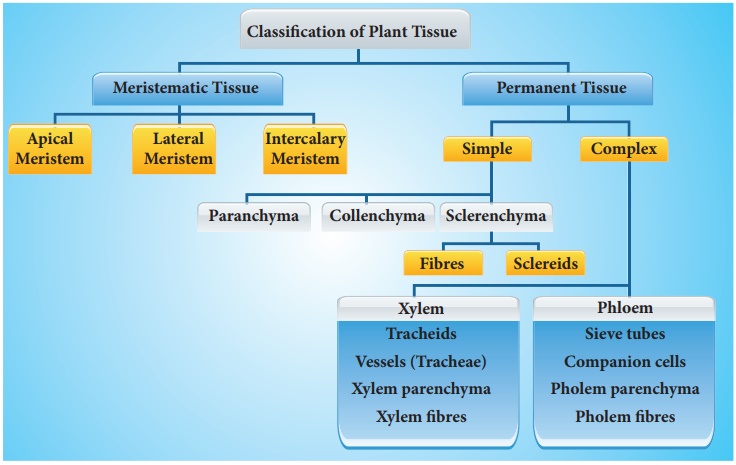
The tissues are classified into two main groups, namely, meristematic and permanent tissues based on whether the cells being formed are capable of dividing or not.
A. Meristematic Tissues
These tissues consist of cells that retain the power of division.
The protoplasm within the cell is dense, the vacuole is smaller or absent.
These cells are isodiametric, without intercellular spaces.
The nucleus is bigger in size.
These cells have thin cellulosic cell wall.
Metabolically active cells with high surface area per unit volume and nucleo-cytoplasmic ratio.
Ergastic substances are absent.
Colourless proplastids are present in cells.
Concept Builder
Classification of meristems :
(1) On the basis of origin and development
(a) Promeristems (Primordial meristem). A group of cells which represent primary stages of meristematic cells. They are represented by few cells found at the apices of shoots and roots. They give rise to primary meristems.
(b) Primary meristems. They originate from promeristems. They are found below the promeristem at shoot and root apices, at the apex of leaves and in intercalary parts. They give rise to primary permanent tissues after differentiation.
(c) Secondary meristems. They are not present from the beginning of the formation of an organ but develop at a later stage. They give rise to secondary permanent tissues. They develop from primary permanent tissue as a result of dedifferentiation, e.g., interfascicular cambium, cork cambium and cambium in dicot roots.
(2) On the basis of position
(a) Apical meristem. These cells or tissues are found at the apices of stem and root. Due to their continuous division the root and stem increase in length. The apical meristem helps the plants to grow in length.
(b) Intercalary meristem. These tissues are intercalated between permanent tissues. These are actually a part of the apical meristem which gets separated from it during the growth of stem in length. The most characteristic example is the stem of grasses and Equisetum. They are responsible for increase in the length of the stems of grasses especially. They are commonly located at the base of the leaves, above the nodes (e.g. grasses) or below the nodes (e.g. mint).
(c) Lateral meristem.These meristems are present along the lateral side of stem and roots. They divide in tangential plane, giving rise to the secondary permanent tissues to the inside and outside and lead to the increase in thickness or girth of the plant body, e.g., intrafascicular cambium , interfascicular cambium and cork cambium.
(3) On the basis of plane of cell division
(a) Rib or file meristem. The cells divide anticlinally in one plane, so row or column of cells is formed, e.g., formation of lateral roots.
(b) Plate meristem. The cells divide anticlinally in two planes, so plate like area is increased, e.g., formation of epidermis and lamina of leaves.
(c) Mass meristem. The cells divide anticlinally in all planes, so that a mass of cells is formed, e.g., formation of spores, cortex, pith, endosperm.
(4) On the basis of function
(a) Protoderm. These are outermost meristematic cells. They form skin or epidermis of plant and epidermal tissue system.
(b) Procambium. These are innermost meristematic cells. They form primary xylem, primary phloem and cambium.
(c) Ground meristem. They form ground or fundamental tissue such as hypodermis, cortex, pith, pericycle, etc.
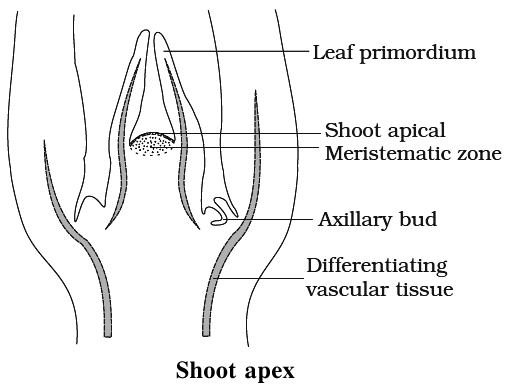
Shoot Apex Organisation
Shoot apex is present immediatley above the youngest leaf primordia. It consists of meristematic cells. Lateral branches of stem and leaves are formed by the activity of shoot apex.
Concept Builder
Many theories have been given to explain shoot apex organisation, such as
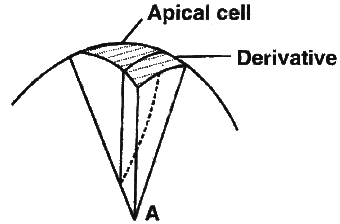
(i) Apical cell theory. It was proposed by Hofmeister and Nageli. According to this theory a single apical cell leads to the development of entire plant body. This theory is applicable to algae, as well as to most of the bryophytes and pteridophytes.
(ii) Histogen theory. It was proposed 'by Hanstein. According to this theory shoot apex consists of following histogens
(a) Dermatogen. Outermost layer. It forms epidermis (skin) and epidermal tissue system.
(b) Periblem. It gives rise to the tissues between epidermis and stele, i.e., cortex and endodermis.
(c) Plerome. Innermost layer. The central mass of cells which gives rise to central stele.

(iii) Tunica-Corpus theory. It was proposed by Schmidt (1924). It is based on plane of division of cells. According to this theory, shoot apex consists of two distinct layers as
(a) Tunica. It is mostly single layered and forms epidermis. The cells of tunica are smaller than corpus and divide by anticlinal divisions mostly.
(b) Corpus. It represents central core with larger cells. The cells divide in all planes (anticlinal and periclinal) .
Sometimes, tunica is multilayered, then only outer layer forms epidermis and the remaining layers with corpus form cortex of shoot.

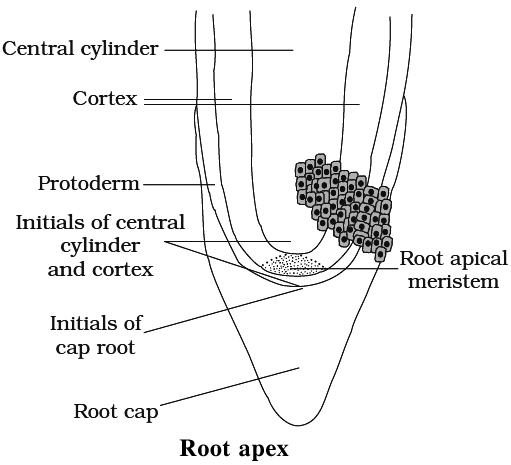
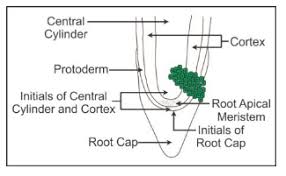
Root Apex Organisation
Root apex consists of mass of meristematic cells.
Root apex is not responsible for the formation of lateral roots.
Root cap is present due to which root meristem becomes subterminal in position.
If root cap is independent in origin, it arises from the calyptrogen.
Concept Builder
Regarding the organisation of root apex, following theories have been put forward.
(i) Korper-Kappe theory. It was proposed by Schuepp (1917). This theory is comparable with the tunica and corpus theory of shoot apex because it is also based upon plane of division. Korper means body and Kappe means cap.
(ii) Quiescent centre theory. It was proposed by Clowes (1956-58) in maize. According to this theory, root apex consists of an inverted cup like structure called the quiescent centre. The cells of this region show very low mitotic activity (quiescent) . They have low amount of RNA, DNA and protein. They are surrounded by layer of actively dividing cells which are responsible for formation of different structures of root. These cells divide only when the root apex gets injured.
B. Permanent Tissues
They are composed of living or dead cells which are derived from the meristematic tissue but have lost their ability to divide.
They are primary permanent tissues, if they are derived from apical or intercalary meristem.
They are secondary permanent tissues if they are derived from lateral meristem.
Permanent tissues are mature cells with permanent special structure and function.
These are of three types:
(I) Simple Tissues
(II) Complex Tissues and
(III) Secretory Tissues
(I) Simple tissues
They are made up of one kind of cells performing similar function.
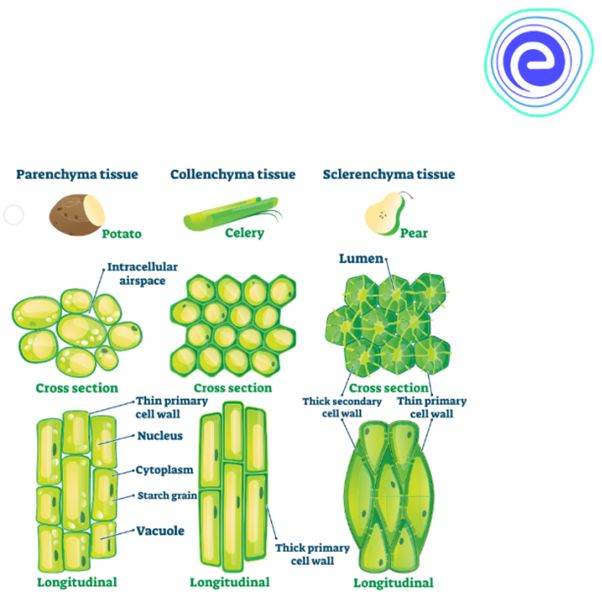
(a) Parenchyma
These cells are found almost in all parts of plants such as roots, stem, leaves, fruits and seeds.
These cells are isodiametric, spherical, oval or polygonal with intercellular spaces.
These cells are living with thin cellulosic cell wall.
Elongated parenchyma with tapering ends is called prosenchyma.
Functions:
(i) The main function is storage of food , e.g., fruits and endosperm.
(ii) Storage of water in fleshy stem and leaves e.g., Opuntia.
(iii) Sometimes, they store secretory substances (ergastic substances) such as tannins, resins and gums and they are called as idioblasts.
(iv) In hydrophytes, they have large intercellular spaces filled with air and are called aerenchyma. They help in circulation of air as well as provide buoyancy to plants.
(v) Sometimes, parenchymatous cells have chloroplast to help in photosynthesis and are called chlorenchyma.
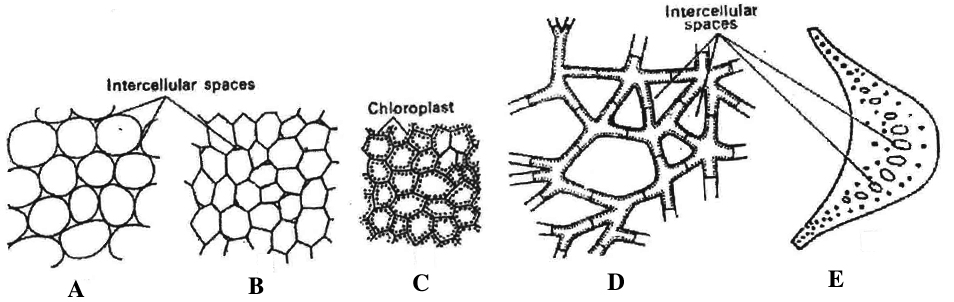
D & E. Aerenchyma
(b) Collenchyma
The term collenchyma was coined by Schleiden.
These cells have thickenings on the cell wall and in corners of intercellular spaces.
It is living mechanical tissue.
They are not found in roots and monocot stems.
These cells form hypodermis in stem and petiol.
The thickening material, in the cell wall contains high amount of pectin and cellulose.
Lignin is absent.
Concept Builder
Collenchyma is of three types :
1. Angular collenchyma - Angular walls thickened e.g., stem of Marigold, Tomato, Datura.
2. Lamellate collenchyma - Tangential walls thickened e.g., stem of Sunflower.
3. Lacunate collenchyma - Lacunate thickening, intercellular spaces are present e.g., stem of Calotropis.
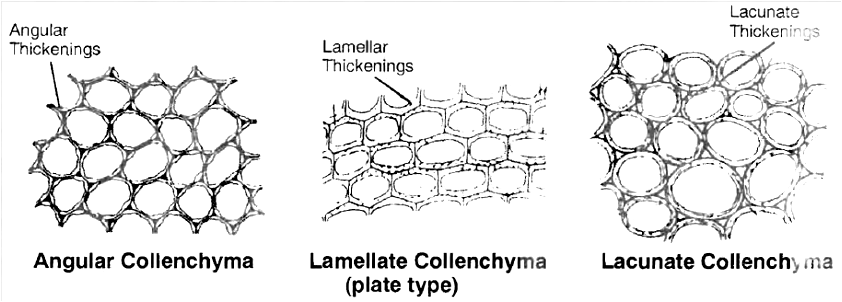
Function :
It provide mechanical support, flexibility and elasticity to the organs and due to peripheral position in stems they resist bending and pulling action of wind . It is especially useful for young plants and herbaceous organs where it is an important supporting tissue.
(c) Sclerenchyma. These cells have thickened secondary walls due to deposition of lignin. On maturity they become dead. These cells have simple pits. They are of two types:
(i) Sclereids
They may be spherical, oval and cylindrical.
They are lignified and extremely thick walled. So the lumen of the cells is almost obliterated.
They are found in hard parts of the plant.
1. Brachysclereids (Stone cells). They are isodiametric and they are found in bark, pith, phloem, cortex, hard endocarp and fleshy portion of some fruits. The grittiness of the fruits like guava and pear is due to these.
2. Macrosclereids (Rod cells). These are elongated, rod like and found in seed coat of leguminous plants.
3. Osteosclereids (Prop cells). These are rod like with dilated ends or barrel shaped e.g., leaves and seed coat of many monocots and seed coat subepidermis in legumes.
4. Asterosclereids (Star cells). These are stellate in form or star shaped. They are common in petiole of floating hydrophytes, like Nymphaea, Lotus and leaves of tea.
5. Trichosclereids (Internal hair). Long, hair like branched sclereids. They are common in hydrophytes and aerial roots of Monstera.
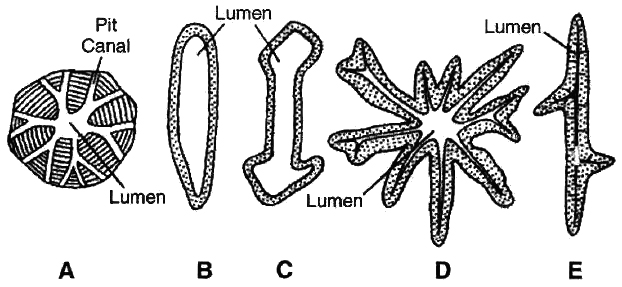
C. Osteosclereid; D. Asterosclereid; E. Trichosclereid
(ii) Sclerenchymatous fibres
They are long with tapering ends.
These are thick-walled cells (lignified).
The fully developed fibre cells are always dead.
The length of fibre varies from 2-550 mm in angiosperms and 1 to 12 mm in gymnosperms.
The fibres are present in hypodermis of monocot stem, in pericycle of many dicots, in secondary wood and in vascular bundle sheath in monocot stems. e.g., Jute, Flax, Hemp etc.
Living fibres are found in Tamarix.
Function: The main function of sclerenchyma is to provide mechanical strength.
(II) Complex tissues
They are made up of different types of cells working as a unit to perform a common function. It includes xylem and phloem.
A. Xylem (by Nageli) or Hadrome (by Haberlandt).
It is chief water conducting element and also provides mechanical strength.
On the basis of origin, xylem is of two types:
(i) Primary xylem
It is derived from procabium during the formation of primary plant body.
It is differentiated into protoxylem (first formed and consists of tracheary elements and xylem parenchyma) and metaxylem (later formed and consists of tracheary elements, xylem parenchyma and fibre).
The cells of metaxylem are bigger in size than protoxylem.
(ii) Secondary xylem.
It is formed from cambium during secondary growth. It is well differentiated into two systems.
(a) Axial or vertical system
1. Tracheary element (Tracheids and Vessels) For conduction of water
2. Xylem or wood fibre For support
3. Xylem parenchyma For storage of food
(b) Ray or horizontal system
Ray parenchyma For storage of food.
c and e. -Xylem parenchyma, d. Wood fibres wood sclerenchyma)
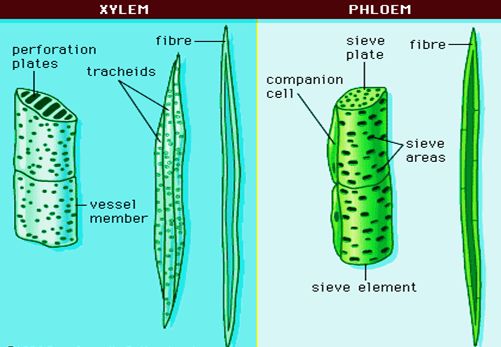
Xylem consists of following types of cells:
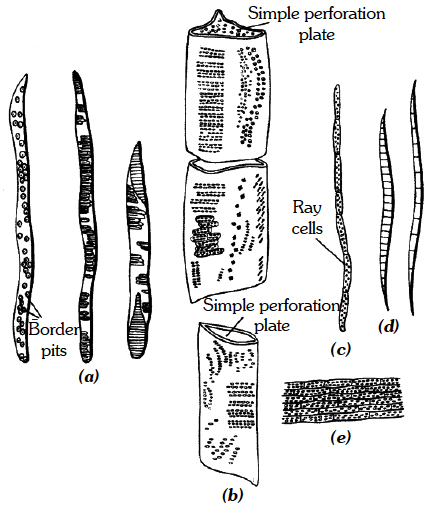
(i) Tracheids.
They are elongated cells with pointed chisel like ends, having no perforations.
Their wall is tough, thickened, lignified and thickening may be annular, spiral, reticulate, scalariform or pitted.
Cells are dead at maturity and have bordered pits.
In pteridophytes and gymnosperms, wood mainly consists of tracheids (no vessels).
In angiosperms, tracheids are associated with vessels.
The main function is conduction of water.
The tracheids are most primitive type of conducting elements in xylem.
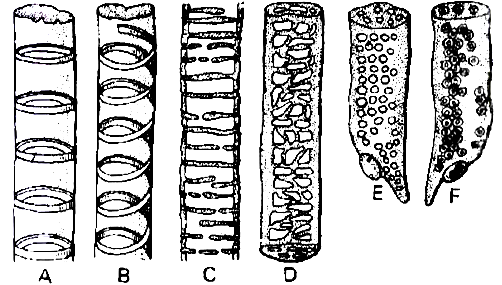
A. Annular B. Spiral C. Scalariform D. Reticulate E. & F. Pitted
(ii) Vessels
They are also elongated and tube like, formed from a row of cells placed end to end.
The partition walls are either perforated or disappear altogether, resulting in an elongated tube.
Walls are thickened and lignified, may have annular, spiral, reticulate or scalariform thickenings.
Vessels are dead at maturity and without nuclei.
The main function is conduction of water.
Vessels are advanced type of conducting elements.
Concept Builder
In pteridophytes and gymnosperms, vessels are absent (non porous wood).
Sometimes, primitive vessels are present in Gnetum and Ephedra (Gnetales).
Vessels are characteristic of angiosperms (porous wood), but they are absent in members of vesselless families like Winteraceae, Trochodendraceae and Tetracentraceae.
(iii) Wood or xylem fibre.
These cells are elongated and pointed at both the ends.
Lumen is completely obliterated.
Cell wall is highly lignified with simple pits.
They are commonly found in secondary xylem.
They may be :
(a) Fibre tracheids. Fibre like tracheids with bordered pits.
(b) Libriform fibre. They have extremely thick walls and simple pits. They provide mechanical support.
(iv) Wood or xylem parenchyma.
They are living parenchymatous cells associated with xylem.
They may occur as axial parenchyma or ray parenchyma.
Concept Builder
When parenchyma is diffused or not associated with vessels, it is called apotracheal parenchyma and when parenchyma surrounds or is associated with vessels, it is called paratracheal parenchyma.
B. Phloem (by Nageli) or Bast or Leptome (by Haberlandt)
It is the main food conducting tissue.
Types of phloem
(a) On the basis of position
(1) External phloem. It is of normal type and is present outside the xylem, e.g., most angiosperms and gymnosperms.
(2) Internal or intraxylary phloem. It originates from procambium and is the primary phloem which occurs on innerside of primary xylem in bicollateral bundles. e.g., Members of Apocyanaceae, Asclepiadaceae, Convolvulaceae, Solanaceae and Cucurbitaceae.
(3) Included or interxylary phloem. It originates from cambium and is secondary phloem which occurs in groups within the secondary xylem, e.g., Leptadaenia, Salvadora, Chenopodium, Boerhaavia, Amaranthus.
(b) On the basis of origin it is of two types:
(1) Primary phloem. It develops from procambium. It does not have radial differentiation or rays are absent. It is differentiated into protophloem (consists of sieve elements and parenchyma) and metaphloem (develops after protophloem and consists of sieve elements, parenchyma and fibre). During the primary growth the protophloem elements are crushed by the surrounding tissues and disappear. This process is known as obliteration.
(2) Secondary phloem. It develops from cambium during secondary growth. It shows radial differentiation. It consists of two distinct systems:
(a) Axial or Vertical system
1. Sieve elements (Sieve tube and companion cells) : For conduction of food
2. Bast fibre : For support
3. Bast Parenchyma: For storage of food
(b) Ray or Horizontal system. Consists of ray parenchyma for storage of food.
Phloem consists of following types of cells
(1) Sieve element.
The sieve elements in angiosperms are sieve tubes which are cylindrical tube like cells with perforated cross walls called sieve plate.
Sieve tubes are associated with companion cells and they are without nuclei.
In pteridophytes and gymnosperms the sieve elements have sieve plates on their lateral walls and are called sieve cells and companion cells are absent.
The walls of sieve tube elements are made up of cellulose and pectic substances.
The cytoplasm is confined to a thin peripheral layer.
P-proteins (Phloem protein) are proteinaceous structures present in sieve tubes and are believed to be responsible for (i) movement of materials and (ii) sealing of pores after wounding.
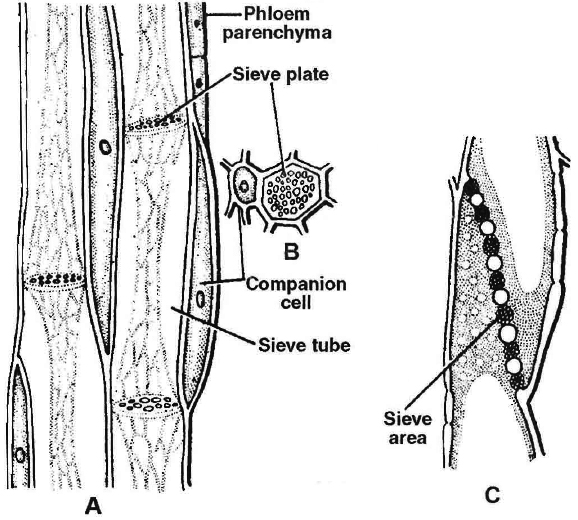
C. L.S. of sieve plate
At the end of growing season a callose plug (made of callose carbohydrate) is deposited on sieve plate in old sieve tubes which inhibits the activity of sieve tubes.
In spring season the callose plug gets dissolved.
(2) Companion cells.
They are elongated, living, parenchymatous, thin walled cells.
They are associated laterally to sieve tubes and have dense cytoplasm and nuclei.
Companion cells are absent in pteridophytes and gymnosperms.
Both sieve tubes and companion cells are related ontogenetically because both develop from the same mother cell.
So, these are called sister cells.
(3) Phloem or bast fibre.
They are absent or fewer in primary phloem and abundantly found in secondary phloem.
They are sclerenchymatous and unbranched fibres associated with phloem.
Phloem fibres of plants like jute, flax and hemp are retted in water and extracted for making ropes and coarse textiles.
(4) Phloem parenchyma.
They are parenchymatous living cells with cellulosic cell wall and nucleus.
The main function is storage of food.
They are not found in monocotyledonous plants.
(III) Secretory Tissues ¬
These tissues perform special function in the plants e.g., secretion of resin, gum, oil and latex.
Concept Builder
These tissues are of two types :
(1) Laticiferous tissues.
They are made up of thin walled, elongated, branched and multinucleate (coenocytic) structures that contain colourless, milky or yellow coloured heterogenous substance called latex.
These are irregularly distributed in the mass of parenchymatous cells.
These tissues are of two types¬ –
(a) Latex cells. They do not fuse and do not form network. Such tissues are called simple or non-articulated laticifers e.g., Calotropis (Asclepiadaceae), Nerium, Vinca (Apocyanaceae), Euphorbia (Euphorbiaceae), Ficus (Moraceae).
(b) Latex vessels. They are formed due to fusion of cells and form network like structure. Such tissues are called compound or articulated laticifers, e.g., Argemone Papaver (Papaveraceae), Sonchus (Compositae), Hevea (rubber plant), Manihot (Euphorbiaceae).
The latex of some plants is of great commercial importance such as¬
(i) Source of commercial rubber is latex of Hevea brasiliensis, Ficus elastica, Cryptostegia, Manihot glaziovii.
(ii) Source of chewing or chickle gum is latex of Achras sapota.
(iii) Source of enzyme papain is latex of Carica papaya. .
(iv) Source of alkaloid opium is latex from immature capsules of Papaver somniferum (Poppy)
(2) Glandular tissues.
They include different types of glands which secrete oils, gums, mucilage, tannins and resins.
They may be :
(a) External glands. They generally occur on the epidermis of stem and leaves e.g., glandular hair in Utricularia, nectar secreting glands in flowers, digestive enzyme secreting glands) in Drosera, Nepenthes (insectivorous plants).
(b) Internal glands. These are present internally, e.g., oil glands in Citrus and Eucalyptus, resinous ducts in Pinus (schizogenous origin) and mucilage secreting glands in leaves, of Piper betel. The glands which secrete essential oils are called osmophores.
The Tissues
Chapter 6
Anatomy of flowering plants
Both plants and animals have structural similarities and differences in their outward appearance that can be easily viewed. Furthermore, the interior structure of living creatures shares certain characteristics. The internal structure and organization of higher plants are discussed in this chapter. Anatomy is the study of a plant's interior structure. Cells are the basic unit of plants, and cells are organized into tissues, which are then organized into organs. Different organs of a plant have different internal structures. Monocots and dicots are considered to be physically distinct within angiosperms. Internal structures also exhibit environmental adaptations.
Tissues
A tissue is a collection of cells that share the same origin and typically execute the same function. Different types of tissues make up a plant. Tissues are divided into two categories: meristematic and permanent tissues, depending on whether or not the cells forming them are capable of dividing.
A. Meristematic Tissues
Meristematic tissue is a type of plant tissue that can divide continuously throughout its existence. Plant growth is generally restricted to meristems, which are specialized regions of active cell division. Meristems come in several forms in plants. Primary meristems include apical and intercalary meristems, which arise early in a plant's life and contribute to the production of the primary plant body.
Apical meristems are meristems that form at the terminals of roots and shoots and create primary tissues. The root apical meristem is found at the root's tip, while the shoot apical meristem is found at the far end of the stem axis. The axillary bud is formed by cells left over from the shoot apical meristem after the production of leaves and stem elongation. These buds can develop a branch or a flower and can be found in the axils of leaves.
Intercalary meristem is a type of meristem that arises between mature tissues. They are present in grasses and restore sections of the plant that have been eaten by grazing herbivores. They are found in the leaves and internodes. These contribute to the internode's lengthening. It is also present in monocots, and pine trees. It contributes to the plant's height.
The secondary or lateral meristem is found in the mature portions of roots and shoots of many plants, especially those that generate woody axes and develop later than the primary meristem. They are meristems that are cylindrical. Lateral meristems include the fascicular vascular cambium, interfascicular cambium, and cork-cambium. The secondary tissues are produced by these cells. Following cell division in both primary and secondary meristems, newly produced cells become architecturally and functionally specialized, and their ability to divide is lost. Permanent or mature cells are the cells that make up the permanent tissues. Specific parts of the apical meristem produce dermal tissues, ground tissues, and vascular tissues during the creation of the main plant body.

B. Permanent Tissues:
Permanent tissues are made up of mature cells that have lost their ability to divide and have taken on a permanent shape, size, and function as a result of meristematic tissue development and differentiation. These tissues' cells can be alive or dead, thin-walled or thick-walled. Permanent tissue cells do not usually divide any further. They are divided into simple tissues and complex tissues.
Simple tissues
are permanent tissues with all cells having the same structure and function.Only one sort of cell makes up a basic tissue. Plants have parenchyma, collenchyma, and sclerenchyma as simple tissues. The parenchyma cells form a majority of the living cells in the plant. The parenchyma's cells are usually isodiametric. They come in a variety of shapes, including spherical, oval, round, polygonal, and elongated. Their cellulose-based walls are quite thin. They might be tightly packed or with minor intercellular gaps. Photosynthesis, storage, and secretion are all processes performed by the parenchyma. In most dicotyledonous plants, the collenchyma is found in layers beneath the epidermis. It can be found as a single layer or in patches. It is made up of cells that have thickened at the corners due to cellulose, hemicellulose, and pectin deposition.Collenchymatous cells can be oval, round, or polygonal, and chloroplasts are common. When chloroplasts are present, these cells digest food. There are no intercellular spaces. They offer mechanical support to the plant's growth portions, such as the young stem and leaf petiole. Long, narrow cells with thick, lignified cell walls and a few or many pits make up sclerenchyma. They are frequently devoid of protoplasts and lifeless. Sclerenchyma can be classified as fibers or sclereids based on differentforms, structures, origins, and development. Fibers are thick-walled, elongated, cells that can be found in groups throughout the plant. Sclereids are dead cells that are spherical, oval, or cylindrical in shape and have very thin cavities (lumen).These are often found in nut fruit walls, fruit pulp such as guava, pear, and sapota, legume seed coats, and tea leaves. Organs are supported mechanically by sclerenchyma.
Complex tissues
are permanent tissues that contain a variety of cell types.They are made up of multiple cell types that work together as a unit. Plants have complex tissues called xylem and phloem. From the roots to the stem and leaves, the xylem serves as a conduit for water and minerals. It also gives the plant parts mechanical strength. Tracheids, vessels, xylem fibres, and xylem parenchyma are the four types of elements that make up the xylem. The xylem of gymnosperms is devoid of vessels. Tracheids are long, tube-like cells with lignified walls and tapering ends. These have no protoplasm and are dead. The inner layers of the cell walls feature different types of thickenings. Tracheids and vessels are the principal water transportation elements in blooming plants.A vessel is a long cylindrical tube-like structure composed of multiple vessel parts, each with lignified walls and a big central chamber. The vascular cells are also protoplasm-free. Members of the vessel are linked through perforations in their common walls. Angiosperms are characterized by the presence of vessels. The walls of xylem fibers are thicker and the central lumens are destroyed. These can be septate or aseptate in nature. Xylem parenchyma cells are live, thin-walled cells with cellulose-based cell walls. They store food items such as starch or fat, as well as other compounds such as tannins. The ray parenchymatous cells are responsible for water radial conduction.
Protoxylem and metaxylem are the two forms of primary xylem. Protoxylem refers to the first created primary xylem elements, while metaxylem refers to the later formed primary xylem components. The protoxylem is located at the organ's center (pith), while the metaxylem is located near the organ's perimeter. Endarch is the name for this sort of primary xylem. Protoxylem is found on the periphery of roots, while metaxylem is found in the centre. Exarch is the name for this type of primary xylem configuration. Food supplies are transported via phloem from leaves to other sections of the plant. Sieve tube elements, companion cells, phloem parenchyma, and phloem fibres make up phloem in angiosperms. Albuminous cells and sieve cells are found in Gymnosperms. Sieve tubes and companion cells aren't present.
Sieve tube elements are similar to companion cells in that they are long, tube-like structures that are aligned longitudinally. The sieve plates are formed by perforating their end walls in a sieve-like pattern. A developed sieve element has a big vacuole and peripheral cytoplasm but no nucleus. The nucleus of companion cells controls the actions of sieve tubes. Companion cells are specialized parenchymatous cells that are intimately linked to sieve tube elements. Between their common longitudinal walls, pit fields connect the sieve tube elements and companion cells. The partner cells aid in maintaining the sieve tubes' pressure gradient. Elongated, tapered cylindrical cells with rich cytoplasm and nucleus make up phloem parenchyma.The cellulose cell wall has pores through which plasmodesmata connections between cells can be made. Food and other compounds like as resins, latex, and mucilage are stored in the phloem parenchyma. The majority of monocotyledons lack phloem parenchyma. Sclerenchymatous cells make up phloem fibers. The primary phloem is devoid of them, whereas the secondary phloem has them. These are long and unbranched, with sharp, needle-like apices. Phloem fibers have a very thick cell wall. These fibers lose their protoplasm and die as they mature. Protophloem is the first created primary phloem, which has tiny sieve tubes, and metaphloem is the second formed primary phloem, which has larger sieve tubes.
The Tissue System
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
THE TISSUE SYSTEM
In response to division of labour, tissues are classified into three systems:
A. Epidermal tissue system
It consists of epidermis and its associated structures.
The epidermal cells are living, parenchymatous and compactly arranged (without intercellular spaces).
In aerial parts, epidermis is covered by cuticle.
The epidermal cells secrete a waxy substance called cutin, which forms a layer of variable thickness (the cuticle) within and on the outer surface of its all walls.
It helps in reducing the loss of water by evaporation.
Cuticle is absent in roots.
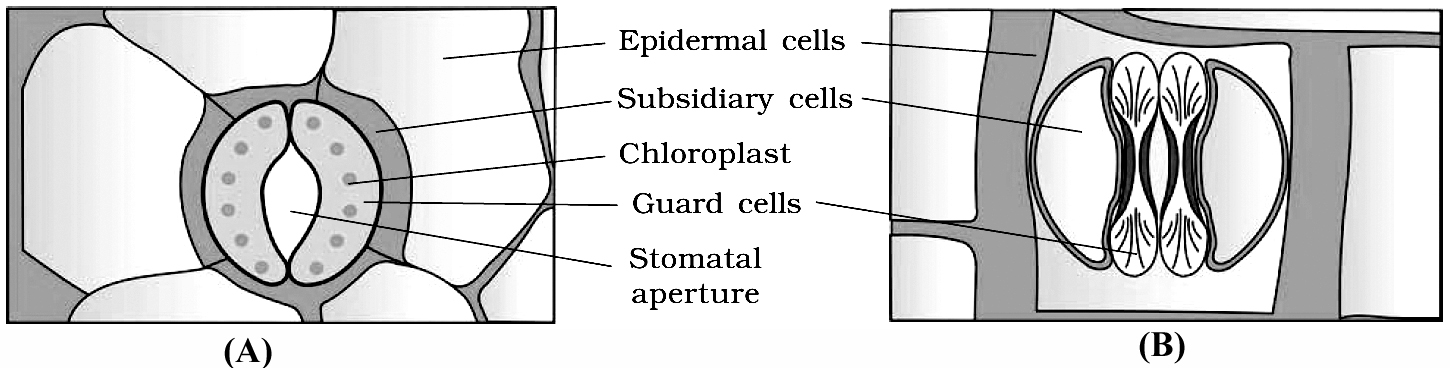
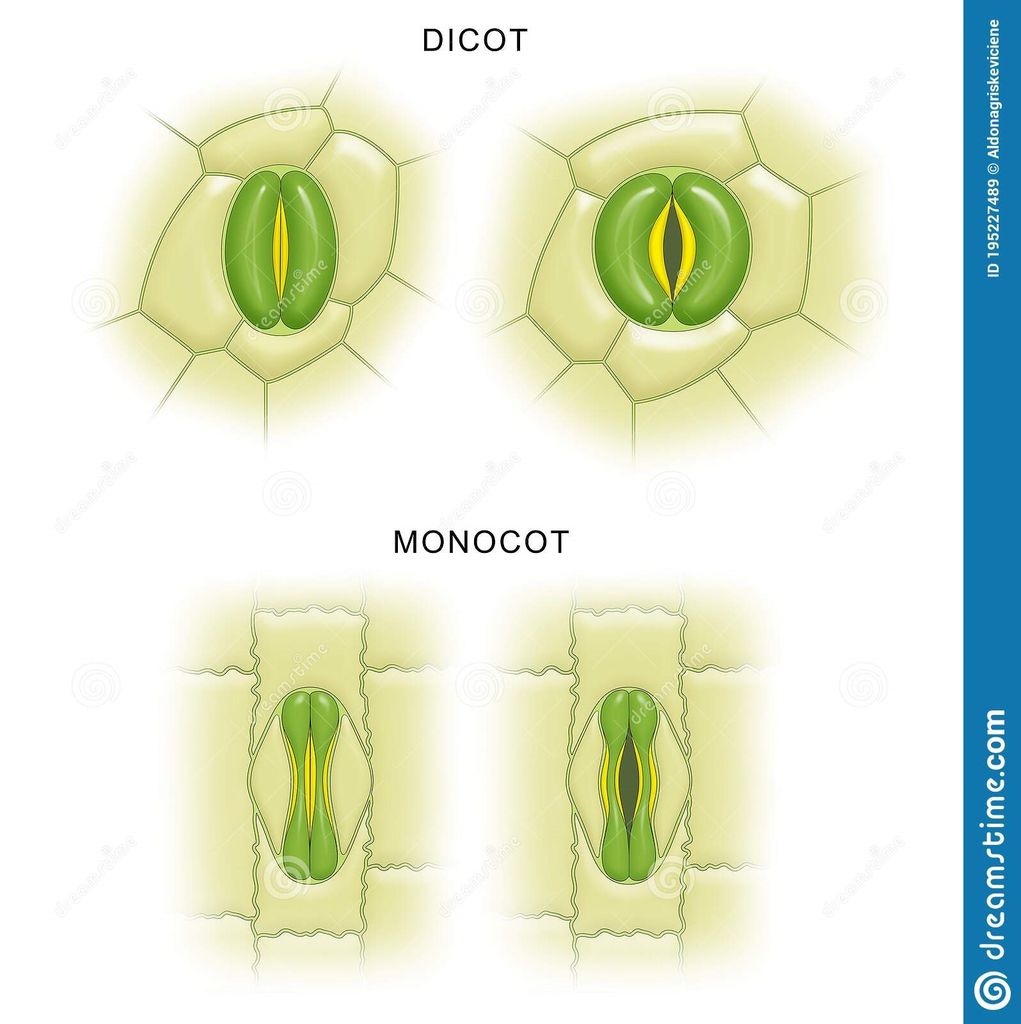
Stomata are structures present in the epidermis of leaves.
Stomata regulate the process of transpiration and gaseous exchange.
Each stoma is composed of two bean shaped cells known as guard cells.
In grasses, the guard cells are dumb-bell shaped.
The outer walls of guard cells (away from the stomatal pore) are thin and the inner walls (towards the stomatal pore) are highly thickened.
The guard cells possess chloroplasts and regulate the opening and closing of stomata.
Sometimes, a few epidermal cells, in the vicinity of the guard cells become specialised in their shape and size and are known as subsidiary cells.
The stomatal aperture, guard cells and the surrounding subsidiary cells are together called stomatal apparatus.
Mostly epidermis is single layered parenchymatous, but is multilayered in leaf of Ficus and Nerium.
Epidermis is mainly protective in nature (external protective tissue).
In grass leaves, motor or bulliform cells are present in upper epidermis.
On stem, the epidermal hairs are called trichomes, which are usually multicellular.
They may be branched or unbranched and soft or stiff. They may even be secretory. These help in preventing water loss due to transpiration.
Concept Builder
In grasses and Equisetum, silica is present in the epidermal cells.
The epidermal cells containing cystoliths are called lithocysts, these are found in Ficus leaves.
B. Ground or fundamental tissue system
It extends from epidermis upto the centre of axis (excluding vascular tissue).
The ground tissue constitutes the following parts :
(a) Cortex. It lies between epidermis and the pericycle. It is further differentiated into
(i) Hypodermis. It is collenchymatous in dicot stem and sclerenchymatous in monocot stem. It provides strength.
(ii) General cortex. It consists of parenchymatous cells. Its main function is storage of food.
(iii) Endodermis (called starch sheath in dicot stem). It is mostly single layered and is made up of parenchymatous, barrel shaped, compactly arranged cells. The inner and radial wall of root endodermis cells have casparian strips. These thick walled endodermal cells are interrupted by thin walled cells just outside the protoxylem patches. These thin walled endodermal cells are called passage cells.
Endodermis behaves as water and air tight dam to check the loss of water and entry of air in xylem elements.
(b) Pericycle. It lies between endodermis and vascular tissue. It is mostly single layered and parenchymatous in roots and sclerenchymatous (mixed with parenchyma) in stem. The pericycle cells just opposite the protoxylem are the seat for the origin of lateral roots. In dicot roots, pericycle form a part of cambium and whole of cork cambium.
(c) Pith. It occupies the central part in dicot stem and monocot root. It is mostly made up of parenchymatous cells. In dicot root, pith is completely crushed by the metaxylem elements. In dicot stem the pith cells between the vascular bundles become radially elongated and are known as primary medullary rays or pith rays. They help in lateral translocation.
C. Vascular tissue system
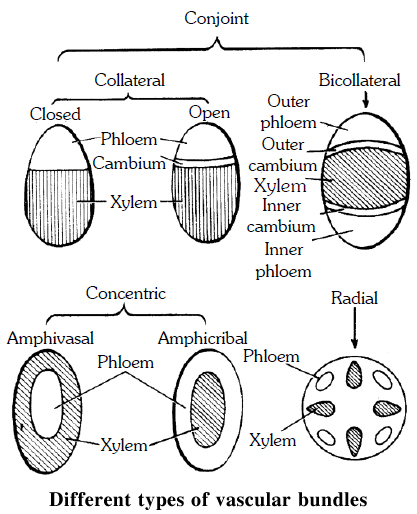
Vascular bundles found in stelar part constitute vascular tissue system.
Xylem, phloem and cambium forms the major part of the vascular bundle.
Vascular bundles may be of following types -
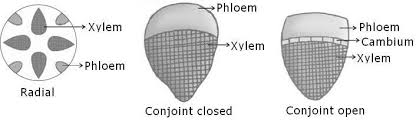
(a) Radial. When the xylem and phloem are arranged on different radii, alternating with each other, e.g., roots.
(b) Conjoint. When xylem and phloem combine in the same bundles and are present on the same radius, e.g., stem. Conjoint vascular bundles may be:
(i) Collateral. Xylem is towards inner side and phloem towards outside.
(ii) Bicollateral. When xylem is surrounded on its both sides by the phloem and cambium e.g., members of Cucurbitaceae and Solanaceae.
• Open. Cambium is present between xylem and phloem, e.g., dicot stem.
• Closed. Cambium is absent between xylem and phloem, e.g., monocot stem.
Concept Builder
Concentric. When one vascular tissue surrounds the other. They are of two types:
(i) Amphicribal or Hadrocentric. The xylem is surrounded on all sides by phloem e.g., ferns.
(ii) Amphivasal or Leptocentric. The phloem is surrounded on all sides by xylem e.g., Yucca, Dracaena.
The Tissue System
The Tissue System
There are three types of tissue systems based on their structure and location.
A. The epidermal tissue system
Epidermal cells, stomata, and epidermal appendages like trichomes and hairs make up the epidermal tissue system, which covers the entire plant body. The epidermis is the major plant body's outermost layer. It is made up of a continuous layer of elongated, compactly packed cells. The epidermis is normally one layer thick. Epidermal cells are parenchymatous, with a big vacuole and a little quantity of cytoplasm along the cell wall. The cuticle, a waxy thick covering on the outside of the epidermis that resists water loss, is commonly present. The roots have no cuticle. The epidermis of leaves has features called stomata. The process of transpiration and gas exchange is regulated by stomata.Guard cells enclose the stomatal pore in each stoma, which are two bean-shaped cells. The guard cells in grasses are formed like a dumbbell. Guard cells have thin outer walls that face away from the stomatal pore and robust interior walls that face the stomatal pore. Guard cells have chloroplasts and control stomata opening and closing. A few epidermal cells near the guard cells can become specialized in shape and size, and these cells are known as subsidiary cells. The Stomatal apparatus is made up of the stomatal orifice, guard cells, and surrounding subsidiary cells. Hairs are found on the epidermis cells.Root hairs are unicellular epidermal elongations that assist absorb water and minerals from the soil. Trichomes are epidermal hairs that grow on the stem. In the shoot system, trichomes are frequently multicellular. They might be soft or stiff, branching or unbranched. They could even be hidden. The trichomes aid in the prevention of water loss by transpiration.
B. The ground tissue system
It is also called the foundational tissue system.The ground tissue is made up of all tissues save the epidermis and vascular bundles. Simple tissues such as parenchyma, collenchyma, and sclerenchyma make up this layer. In the cortex, pericycle, pith, and medullary rays, as well as the primary stems and roots, parenchymatous cells are commonly seen. The mesophyll is the ground tissue in leaves that is made up of thin-walled chloroplast-containing cells. Ground tissue includes parenchyma (photosynthesis in the leaves and storage in the roots), collenchyma (shoot support in areas of active growth), and sclerenchyma (shoot support in areas where growth has ceased) and is the site of photosynthesis, provides a supporting matrix for the vascular tissue, provides structural support for the stem, and helps to store water and sugars, depending on the cell type and location in the plant.
C. The vascular tissue system:
Also called conducting tissue system. The vascular system is made up of two complicated tissues: phloem and xylem. Vascular bundles are made up of the xylem and phloem. Between the phloem and xylem in dicotyledonous stems, there is cambium. Because of the existence of cambium, such vascular bundles have the ability to produce secondary xylem and phloem tissues, and are hence referred to as open vascular bundles. The cambium is absent from the vascular bundles of monocotyledons. As a result, they are referred to as closed since they do not generate additional tissues. Radial arrangement occurs when xylem and phloem within a vascular bundle are placed in an alternative fashion along distinct radii, as in roots.The xylem and phloem are located along the same radius of vascular bundles in conjoint type vascular bundles. In stems and leaves, vascular bundles are common. Phloem is usually exclusively seen on the outer side of the xylem in conjoint vascular bundles.
Anatomy of Dicotyledonous and Monocotyledonous Plants
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
ANATOMY OF DlCOT AND MONOCOT PLANTS
Anatomy of Root
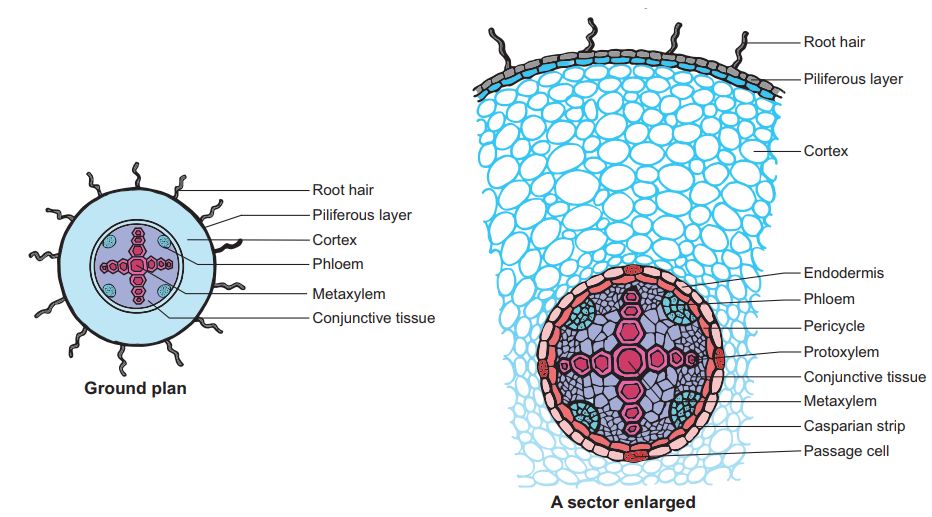
Anatomically, three zones can be distinguished in a root. These are:
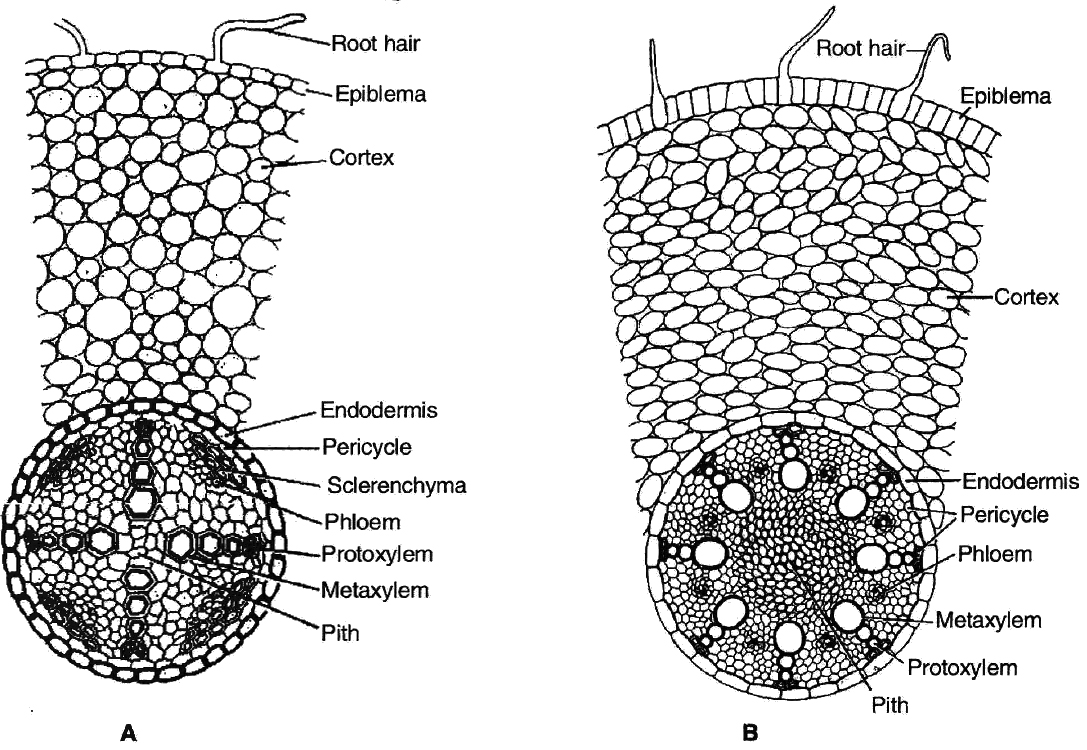
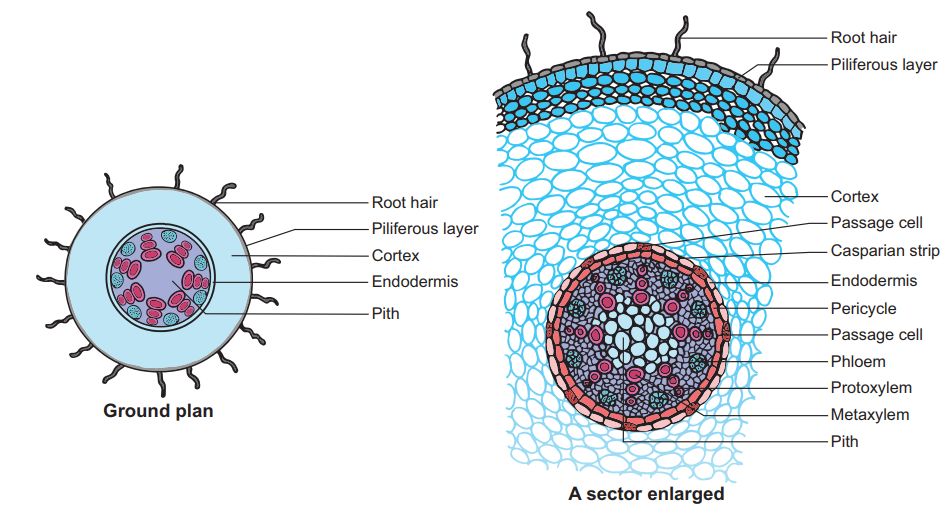
(i) Epidermis. It is single layered (uniseriate) and consists of tightly placed, thin walled, uncutinised cells. This epidermis layer is called as epiblema or rhizodermis. Epiblema in younger roots bears unicellular root hairs (water absorbing organs), and is also called piliferous layer.
(ii) Cortex. It consists of thin walled parenchymatous cells with intercellular spaces. In most monocots and some dicots, the cortex layer below epidermis becomes suberised to form protective tissue called exodermis. The cells of cortex store food material (e.g., carrot). The innermost layer of cortex develops into endodermis. It is made up of closely packed living cells characterised by the presence of band like thickenings made of lignin and suberin on their radial and tangential walls. These bands or strips are called Casparian bands or strips. Some cells of endodermis lying opposite to protoxylem remain thin walled and are called passage cells which allow radial diffusion of water.
(iii) Vascular bundles. Vascular bundles are radial and exarch. The centre of monocot root is occupied by parenchymatous cells called pith.
Differences between dicot and monocot root

Anatomy of Stem
1. Primary structure of dicot stem
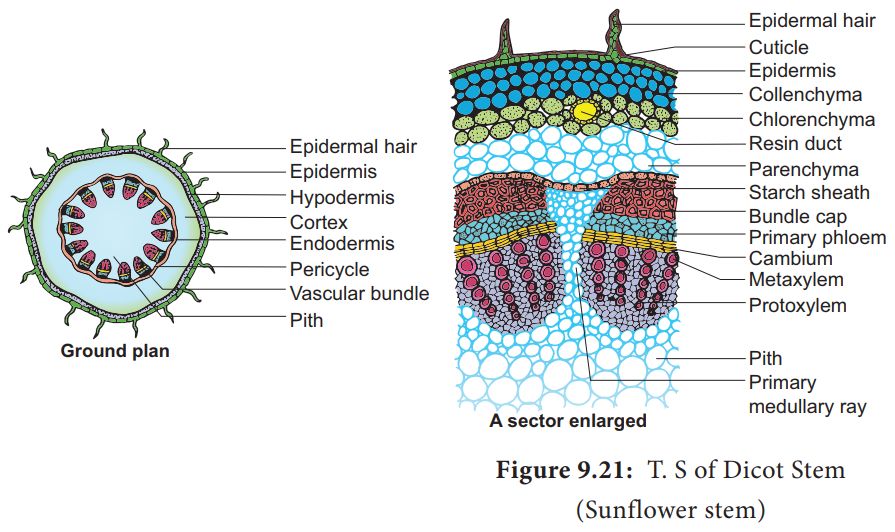
Dicot stem consists of following layers:
(i) Epidermis: It is the outermost layer consisting of single layer of closely arranged cells with cuticle (cutinized). It bears multicellular hairs.
(ii) Cortex: It is differentiated into hypodermis, general cortex and endodermis. Hypodermis is collenchymatous. General cortex is parenchymatous. Endodermis is wavy. It has starch grains hence it is called starch sheath or endodermoid.
(iii) Pericycle: It lies inner to endodermis. Pericycle is few layered thick. Above vascular bundle, it is sclerenchymatous and outside medullary rays it is parenchymatous.
(iv) Vascular bundles: These are in the form of a ring or eustele. They are conjoint, collateral and open. In the family Cucurbitaceae, the stem is wavy, having five ridges and five furrows and vascular bundles are present in ridges and furrows. Vascular bundles are bicollateral and open.
(v) Medullary or pith rays: These are radial strips of parenchyma present between adjacent vascular bundles. They help in radial conduction of food.
(vi) Pith : It is the central portion of stem consisting of parenchymatous cells with intercellular spaces. Narrow, radially elongated parenchymatous cells extend from pith toward the periphery are called medullary rays. The main function is food storage.
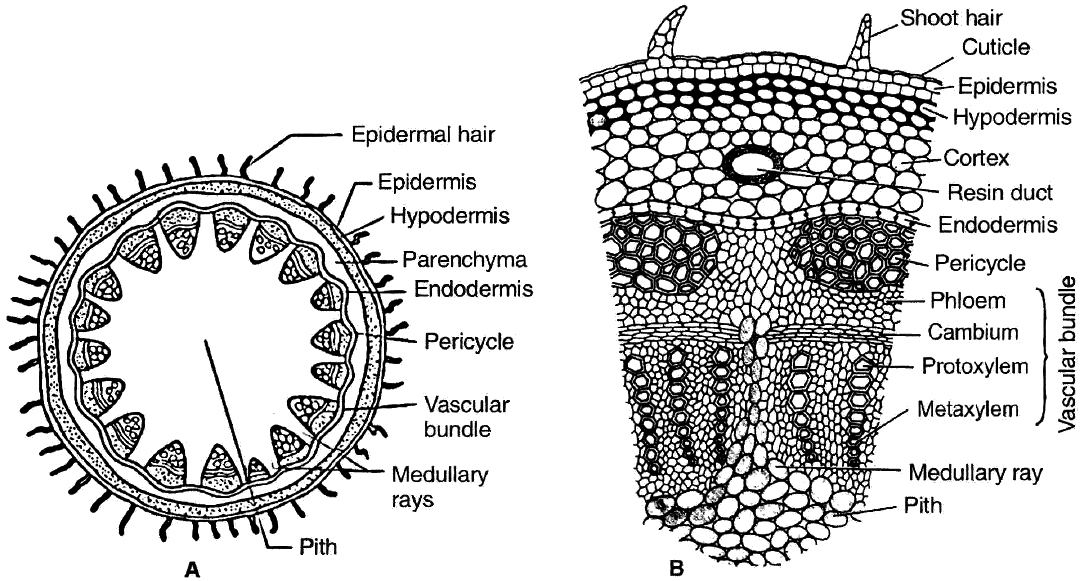
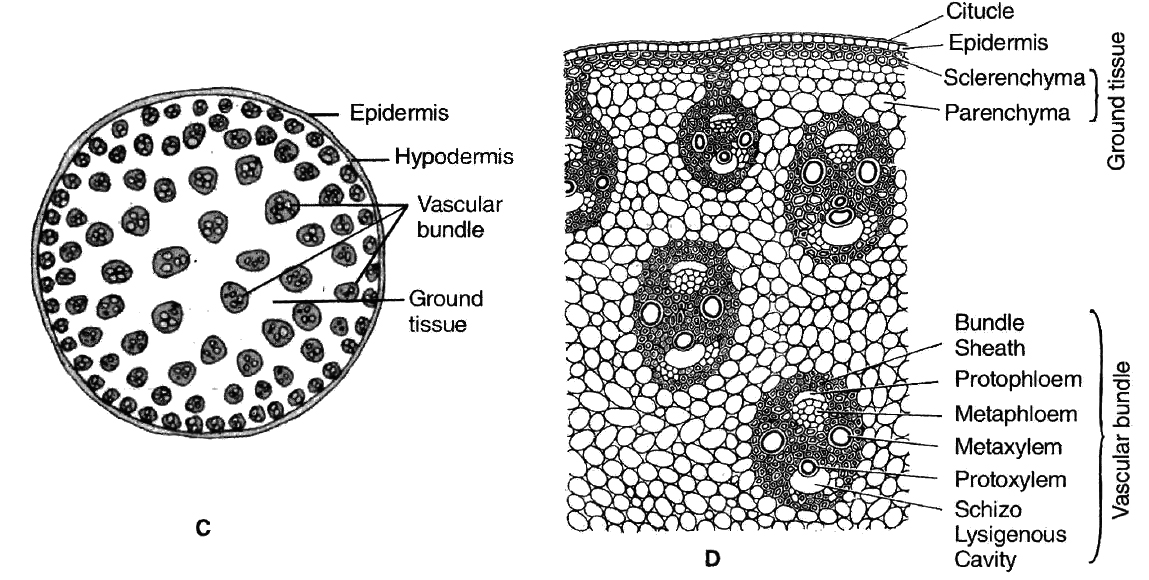
Part of transverse sections of stem: A-B. Dicot stem C-D. Monocot stem
2. Primary structure of monocot stem
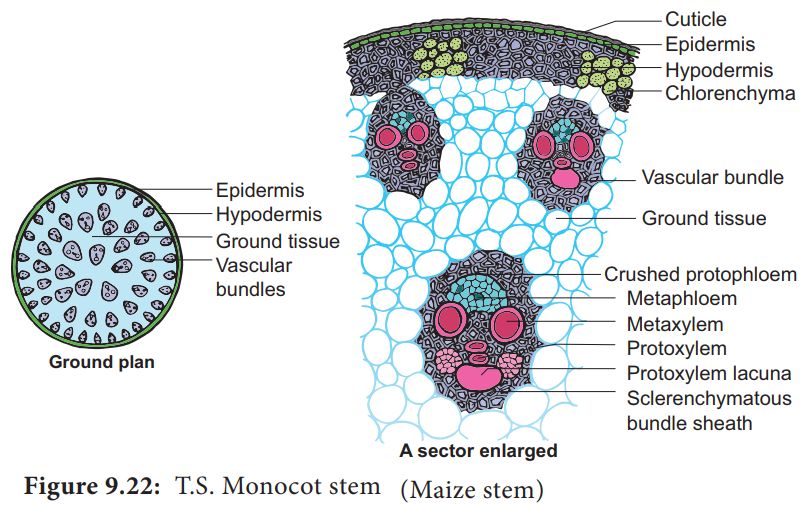
Monocot stem consists of following layers:
(i) Epidermis: It is the outermost layer and consists of compactly arranged parenchyma cells which are usually covered with cuticle.
(ii) Hypodermis: Cells of hypodermis are sclerenchymatous, providing mechanical strength to the stem.
(iii) Ground tissue: All the tissues inner to hypodermis represents the ground tissue. It is made up of parenchymatous cells rich in food reserve, like starch.
(iv) Vascular bundles: They lie scattered in the ground tissue. Each vascular bundle is surrounded by 2 or 3 layered sclerenchymatous sheath called as bundle sheath. The vascular bundles are conjoint, colateral, closed and endarch (Atactostele). Vessels are arranged in V shaped manner. Schizolysigenous water cavity or canals are present below protoxylem.
Differences between dicot and monocot stem anatomy

Anatomy of Leaf
1. Structure of dorsiventral leaf (dicot) :
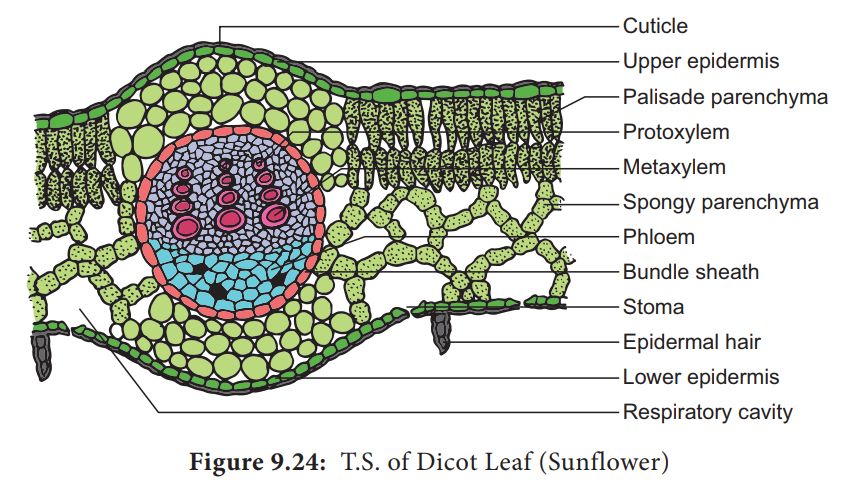
In cross section of dicot leaf, following parts can be observed
(i) Epidermis.
The upper and lower surfaces are covered by the epidermis.
Cells of epidermis are parenchymatous and are closely packed together without any intercellular spaces.
Mostly the stomata are restricted to lower surface of leaf. Such leaves are called hypostomatic.
The outer walls of the epidermal cells are thickened and cutinized which prevents the loss of water.
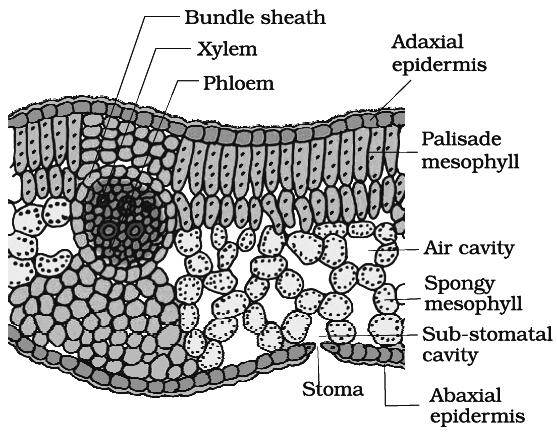
(ii) Mesophyll.
Between the two epidermal layers, there are numerous parenchyma cells which constitute the mesophyll.
In dicots, there are two distinct layers of mesophyll-the palisade (upper layer consisting of closely arranged column shaped cells containing abundant chloroplasts) and spongy tissue (the lower layer of irregularly shaped cells containing fewer chloroplasts).
(iii) Vascular bundles.
Vascular bundles in the leaf are located in the midrib and the veins.
Vascular bundles are conjoint, collateral and closed. Bundles are surrounded by a compact layer of parenchymatous cells which is called bundle sheath.
The xylem (protoxlem) is towards upper epidermis (adaxial) and the phloem on the lower epidermis (abaxial).
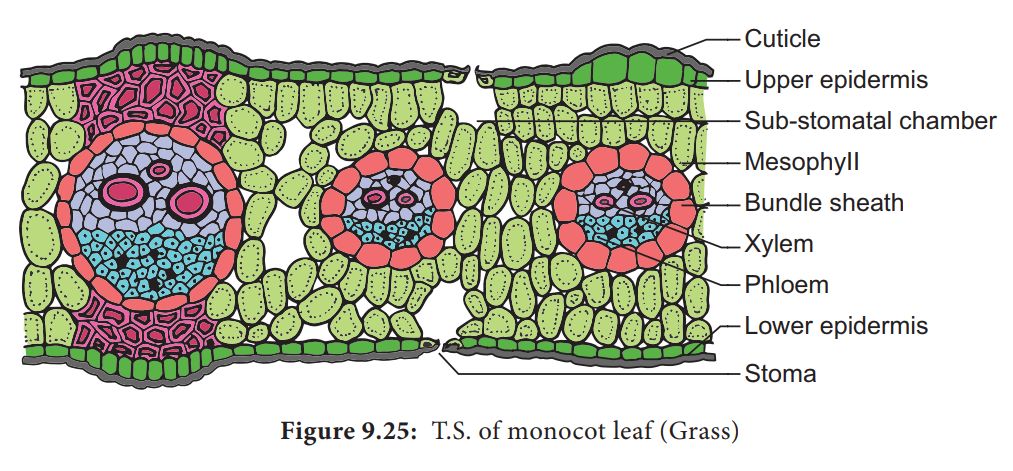
2. Structure of isobilateral leaf (monocot) :
Like the dicot leaf, it can also be differentiated into three types of tissues:
(i) Epidermis.
It consists of upper and lower epidermis, both of which may be interrupted by equal number of stomata.
Both the epidermal layers are cutinized. In some grasses e.g., Poa, Agropyron Maize, Psamma, epidermal cells are large with thin flexible walls which are called motor or bulliform cells.
These cells help in the rolling and unrolling of leaves.
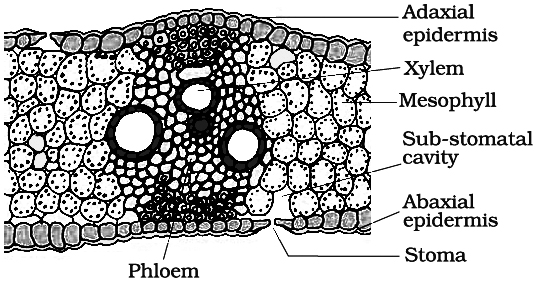
(ii) Mesophyll. Mesophyll cells are not differentiated into palisade and spongy parenchyma. Mesophyll cells are made up of parenchyma cells which have chloroplasts.
(iii) Vascular bundles. They are arranged in parallel manner. Vascular bundles are conjoint, collateral, closed and enclosed by a bundle sheath. The xylem is towards the upper side (adaxial surface) and phloem on the lower side (abaxial surface).
Anatomy of Dicotyledonous and Monocotyledonous Plants
Anatomy of dicotyledonous and monocotyledonous plants
Transverse sections of the mature zones of roots, stems, and leaves are useful for better understanding the tissue organization of these organs.
A.Dicotyledonous Root: The interior tissue organization is as follows:
1. Epiblema is the outermost layer in the interior tissue architecture. Epiblema cells emerge as unicellular root hairs on many occasions.
2. The cortex is made up of many layers of parenchymal cells with thin walls and intercellular gaps.
3. Endodermis is the cortex's deepest layer. There are no intercellular gaps in the single layer of barrel-shaped cells.
4. Water-impermeable waxy substance suberin is deposited in the form of Casparian strips on the tangential as well as radial walls of endodermal cells.
5. A few layers of thick-walled parenchymatous cells called pericycle are found next to the endodermis. During secondary growth, these cells initiate the formation of lateral roots and vascular cambium.
6. The pith is small and undetectable.
7. Conjuctive tissue is made up of parenchymatous cells that reside between the xylem and the phloem. Two to four xylem and phloem patches are typical. A cambium ring forms later between the xylem and the phloem.
8. The stele is made up of all tissues on the inner side of the endodermis, such as the pericycle, vascular bundles, and pith.
B. Monocotyledonous Root: In many ways, the monocot root's anatomy is identical to that of the dicot root.
1. Epidermis, cortex, endodermis, pericycle, vascular bundles, and pith are all present.
2. Unlike the dicot root, which has fewer xylem bundles, the monocot root frequently has more than six (polyarch) xylem bundles.
3. The pith is thick and developed.
4. There is no secondary development in monocotyledonous roots.
C. Dicotyledonous Stem:
1. The epidermis is the stem's outermost protective covering, as shown in this transverse section of a typical immature dicotyledonous stem.
2. It may have trichomes and a few stomata and is covered in a thin layer of cuticle.
3. The cortex is made up of cells organized in many layers between the epidermis and the pericycle. It is divided into three zones.
4. Just below the epidermis, the outer hypodermis consists of a few layers of collenchymatous cells that offer mechanical support to the embryonic stem.
5. The cortical layers underneath the hypodermis are made up of spherical parenchymatous cells with visible intercellular gaps.
6. The endodermis is the deepest layer andbecause the endodermis cells are densely packed with starch grains, the layer is also known as the starch sheath.
7. Pericycle appears as semi-lunar patches of sclerenchyma on the inner side of the endodermis and above the phloem.
8. Medullary rays are a few layers of radially arranged parenchymatous cells that lie between the vascular bundles.
9. A ring of vascular bundles surrounds the stem of a dicot. Each vascular bundle is joined, open, and has a protoxylem at the end.
10. The pith is made up of a large number of spherical, parenchymatous cells with extensive intercellular gaps that occupy the middle region of the stem.
D. Monocotyledonous Stem:
1.A sclerenchymatous hypodermis, numerous distributed vascular bundles, each enclosed by a sclerenchymatous bundle sheath, and a broad, prominent parenchymatous ground tissue characterize the monocot stem.
2. The vascular bundles are joined and closed together. Vascular bundles at the periphery are typically smaller than those in the center.
3. The phloem parenchyma is missing, and there are water-filled holes inside the vascular bundles.
E. Dicotyledonous Leaf:
They are also called Dorsiventral Leaves.
1. The epidermis, mesophyll, and vascular system are visible in a vertical piece of a dorsiventral leaf through the lamina.
2. The epidermis of the leaf has a prominent cuticle that covers both the upper (adaxial epidermis) and bottom (abaxial epidermis).
3. In general, the abaxial epidermis has more stomata than the adaxial epidermis.
4. Stomata may even be absent in the upper epidermis. The mesophyll is the tissue that lies between the top and lower epidermis.
5. The mesophyll is made up of parenchyma, which contains chloroplasts and performs photosynthesis.
6. The palisade parenchyma and the spongy parenchyma are two types of cells. The elongated cells that make up the adaxial palisade hepatocytes are organized vertically and parallel to each other.
7. The spongy parenchyma, which is oval or spherical and loosely organized, is found underneath the palisade cells and reaches the lower epidermis. Between these cells are various huge gaps and air cavities.
8. Vascular bundles can be found in the veins and midrib of the vascular system. The size of the vascular bundles is determined by the vein size.
9. The veins in the dicot leaves' reticulate venation vary in thickness. A layer of thick-walled bundle sheath cells surrounds the vascular bundles.
F. Monocot Leaf:
They are also known as Isobilateral Leaves.In many ways, the anatomy of the isobilateral leaf is identical to that of the dorsiventral leaf. It demonstrates the following distinguishing characteristics.
1. Stomata are present on both epidermal surfaces in an isobilateral leaf, and the mesophyll is not divided into palisade and spongy parenchyma.
2. Certain adaxial epidermal cells along veins in grasses transform into huge, empty, colorless cells called Bulliform cells.
3. The leaf surface is exposed when the bulliform cells in the leaves have absorbed water and are turgid. They curl the leaves inwards to reduce water loss when they are flaccid due to water stress.
4. In vertical sections of monocot leaves, the parallel venation is represented in the near comparable diameters of vascular bundles.
Secondary Growth
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
SECONDARY GROWTH
Increase in girth or thickness or diameter of the axis occurs due to formation of new tissues as a result of joint activity of vascular cambium and cork cambium in steler and extrasteler region respectively.
It occurs in the root and stem of gymnosperms and dicots.
Secondary growth in dicot stem completes in following steps :
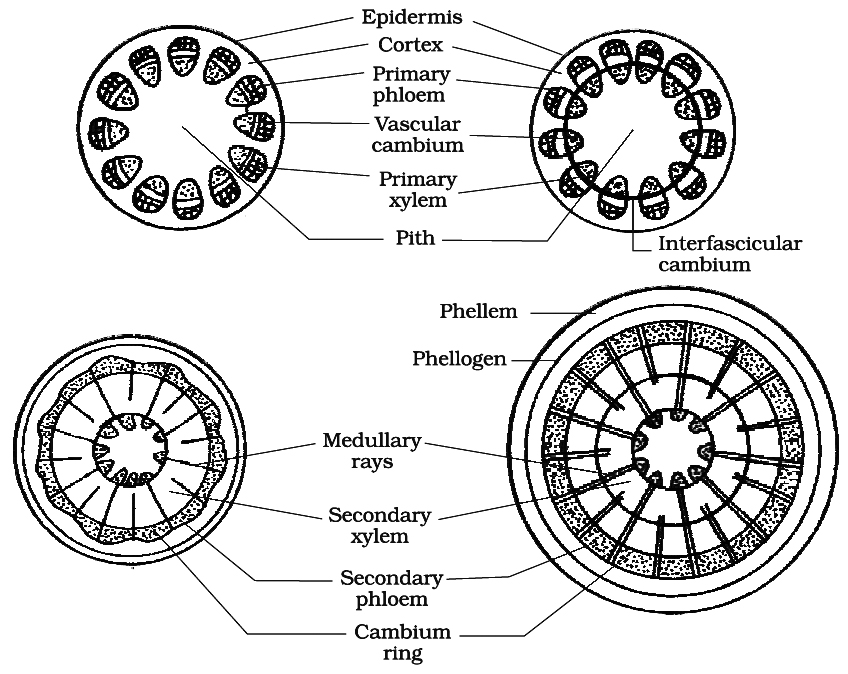
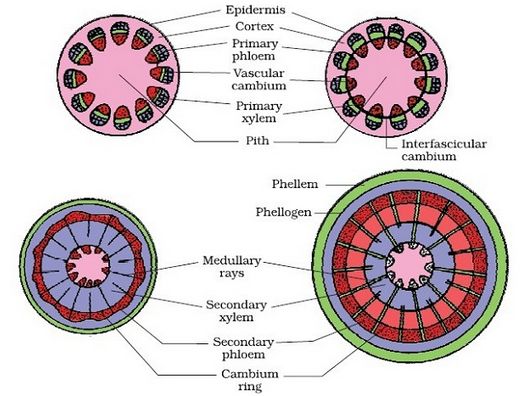
A. Formation of vascular cambium ring :
(i) Intrafascicular cambium : It is primary in origin and is present between primary phloem and primary xylem.
(ii) Interfascicular cambium : It is true secondary meristem. It originates from parenchyma cells of medullary rays region. It lies in between the vascular bundles.
(iii) Vascular cambium ring: Both intrafascicular and interfascicular cambia join together and form a cambium ring.
Cells of cambium are of two types:
(a) Fusiform initials : They form tracheids, vessels, fibres and axial parenchyma in secondary xylem and sieve tubes, companion cells, fibres and axial parenchyma in secondary phloem.
(b) Ray initials : These are isodiametric and form ray parenchyma (vascular rays).
(iv) Periclinal division in cells of vascular cambium ring.
(v) Formation of secondary phloem (outside the vascular cambium) and secondary xylem (inner to vascular cambium). The amount of secondary xylem produced is 8-10 times greater than secondary phloem.
(vi) Due to formation of secondary phloem primary phloem is crushed to death, known as obliteration. Primary xylem being dead and lignified, persists in the pith region by replacing the pith cells.
(vii) At some places, the cambium forms a narrow band of parenchyma, which passes through the secondary xylem and the secondary phloem in the radial directions. These are the secondary medullary rays.
(viii) Formation of secondary structures i.e. annual rings, sapwood and heart wood, hardwood and softwood etc.
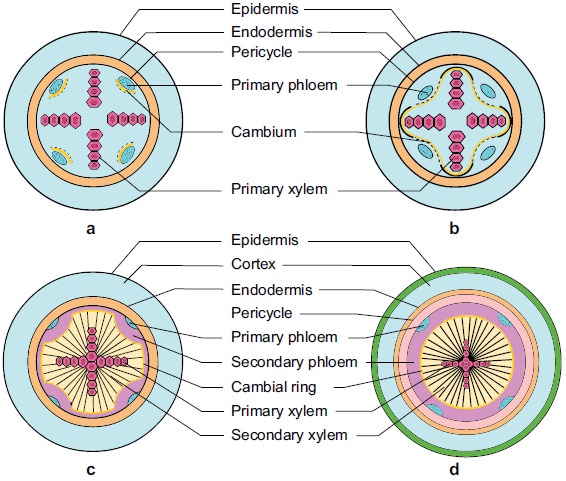
Different stages of secondary growth in a typical dicot stem
(a) Annual rings:
These are formed by the seasonal activity of vascular cambium.
Cambium is not uniformly active throughout the year.
In spring or summer cambium is more active and form large sized xylem elements (vessels) which constitute spring or early wood.
In Autumn or winter, cambium is less active and cuts off small sized xylem elements (vessels) and constitute autumn wood or late wood.
Both autumn and spring wood constitute a growth or annual ring.
In one year only one growth ring is formed.
In successive years numerous growth rings are formed.
Thus by counting the number of annual rings in the main stem at the base we can determine the age of a tree.
This branch of science is known as dendrochronology.
Concept Builder
Growth rings are distinct or sharply demarcated in the plants of temperate climate e.g., Shimla, Nainital, Mussourie due to presence of contrasting seasonal variations.
Growth rings are not distinct or sharply demarcated in the trees of tropical climate (near equator) e.g., Calcutta, Bombay, Madras due to absence of contrasting seasonal variations.
(b) Heart wood and sap wood:
The young elements of secondary xylem in the peripheral region constitute sap wood or alburnum.
It is light in colour and physiologically active.
The water conduction takes place through sap wood.
Sap wood is converted into heart wood or duramen in the central region.
It is dark in colour due to deposition of tannins, gums, resins and is physiologically inactive (almost dead) and provides only mechanical support.
During the conversion of sap wood into heartwood, the most important change is development of tyloses in the heart wood.
Tyloses are balloon like structures in lumen of vessels, developing from xylem parenchyma.
These tyloses block the passage of xylem vessels and are also called tracheal plug.
The heart wood is commercially used as wood.
When the plant is made hollow, it will not die because the water conduction takes place through sap wood.
The heart wood is well developed in Morus alba (Mulberry).
The heart wood is absent in Populus and Salix plant.
The wood of Tectona grandis is termite resistant.
As a tree grows older thickness of heartwood increases and sap wood remains same.
Heart wood is much more durable and resistant to microorganisms, insects and pests etc. than sap wood.
Concept Builder
Wood of dicot trees is called porous or hard wood because it consists of vessels (pores).
The wood of gymnosperms does not contain vessels (pores) and is known as soft or non porous wood.
Such wood consists of 90 to 95% tracheids and 5 to 10% of ray cells.
Sap wood will decay faster if exposed freely to the air.
On the basis of distribution and size of vessels, porous wood is of two types:
(a) Diffuse porous wood (primitive) : Vessels of same size are uniformly distributed throughout the growth, e.g., Pyrus, Betula.
(b) Ring porous wood (advanced) : Large vessels are formed in early wood when the need of water is great and small vessels are formed in late wood, e.g., Quercus, Morus.
B. Formation of cork cambium:
Cork cambium or phellogen develops from outer layer of cortex.
It produces secondary cortex or phelloderm on innerside and cork or phellem on outer side.
The cells of phellem are dead, suberized and impervious to water.
Cork cells are airtight and used as bottle stopper or cork.
The bottle cork is prepared from the cork of Quercus suber (Oak tree).
Cells of phelloderm are thin walled, living and store food. Phellem, phellogen and phelloderm are collectively called periderm.
Periderm is secondary protective tissue.
Due to pressure of secondary tissues, epidermis ruptures and cortex is largely lost after two or three years of secondary growth.
In the cork layer the lenticels are present which are meant for gaseous exchange.
In cork, lenticels have loosely arranged cells called complementary cells with intercellular spaces.
For bottle corks the cork, is processed in such manner, so that lenticels come in vertical direction.
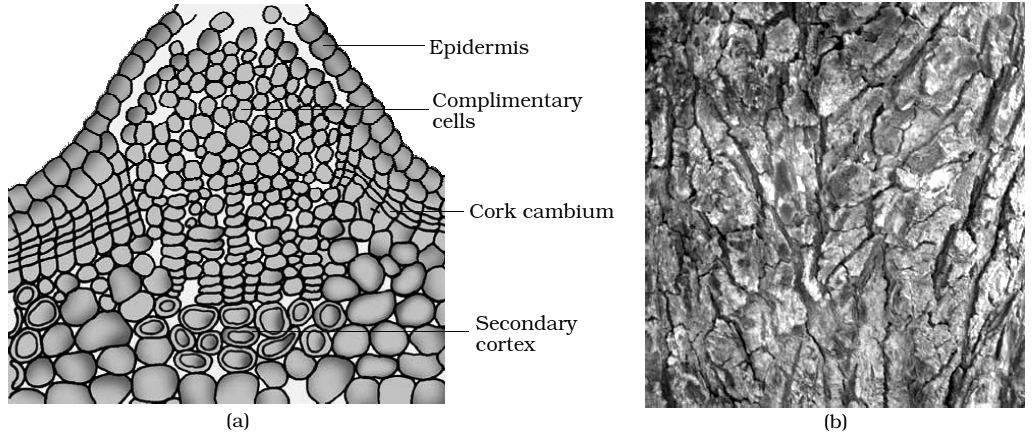
Bark is a non-technical term for all the dead and living tissues outside the vascular cambium. Bark formed early in the season is called soft or early bark. Towards the end of the season late or hard bark is formed.
Concept Builder
(i) Scaly bark: Develops in strips e.g., Acacia, Psidium.
(ii) Ring bark: Develops in the form of sheet or ring, e.g., Betula (Bhojpatra).
The outermost layer of bark is dead and called as rhytidome.
The bark of Betula was being used as substitute of paper in ancient time to write manuscript.
Secondary Growth in Dicot Root
Vascular bundles in dicot root are radial, exarch and mostly triarch.
Vascular cambium is formed secondarily from conjuctive parenchyma cells lying just below each phloem strand.
Thus, the number of cambium strips formed equals the number of phloem strands.
The cells of pericycle lying outside the protoxylem also become meristematic to form part of strips of cambium.
These cambial strips join the first formed cambium strips to form complete, but wavy ring of vascular cambium.
This cambium ring produces secondary xylem on inner side and secondary phloem on outer side.
In roots, the growth rings are not distinct because there is no seasonal variation under the soil.
From the outer layers of pericycle arises the phellogen which cuts phellem (cork) on the outer side and secondary cortex or phelloderm toward the inner side.
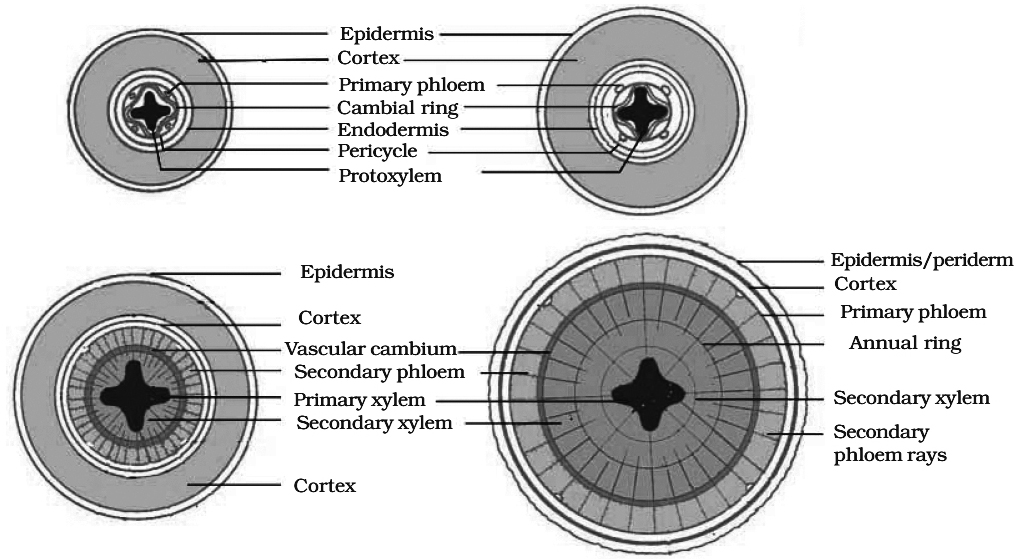
Diagrams showing secondary growth in dicot root
Concept Builder
(i) Dicots with scattered vascular bundles in stem, e.g., Podophyllum, Peperomia, Piper, Papaver.
(ii) Cortical vascular bundles, e.g., Nyctanthus, Kalanchoe, Casuarina.
(iii) Medullary bundles, e.g.; Mirabilis, Bougainvillea, Amaranthus, Achyranthus.
(iv) Polystelic condition, e.g., Primula, Dianthera
(v) Anomalous or abnormal secondary growth occurs in Bougainvillea, Boerhaavia, Chenopodium, Aristolochia.
(vi) Some monocots show abnormal secondary growth by meristematic tissue which develops around vascular bundles, e,g., Dracaena, Yucca, Agave .
(vii) Virgin cork is first formed periderm.
(viii) Wound cork: It is secondary meristem; formed below injured area. It forms cork on outer side and callus below; which heals the wound.
(ix) Abnormal secondary growth in dicot root occurs in Beet root (Beta vulgaris) and Sweet potato (Ipomoea batatas) by the formation of numerous accessory rings of cambium which cut more storage parenchyma in secondary phloem and less secondary xylem.
(x) Homoxylous wood-wood of vesselless dicots, e.g., Ranales (Winteraceae, Tetracentraceae, Trochodendraceae) .
(xi) Heteroxylous wood is wood of vessel bearing dicots.
(xii) Polyderm is a special type of protective tissue occurs in roots and underground stems of members of Rosaceae and Myrtaceae. Its outermost layer is dead and suberized.
Summary
Anatomically, a plant is made of different kinds of tissues. The plant tissues are broadly classified into meristematic (apical, lateral and intercalary) and permanent (simple and complex).
Assimilation of food and its storage, transportation of water, minerals and photosynthates, and mechanical support are the main functions of tissues.
There are three types of tissue systems -epidermal, ground and vascular.
The epidermal tissue systems are made of epidermal cells, stomata and the epidermal appendages.
The ground tissue system forms the main bulk of the plant. It is divided into three zones cortex, pericycle and pith.
The vascular tissue system is formed by the xylem and phloem,
On the basis of presence of cambium, location of xylem and phloem, the vascular bundles are of different types. The vascular bundles form the conducting tissue and trans locate water, minerals and food material.
Monocotyledonous and dicotyledonous plants show marked variation in their internal structures. They differ in type, number and location of vascular, bundles.
The secondary growth occurs in most 6fthe dicotyledonous roots and stems and it increases the girth (diameter) of the organs by the activity of the vascular cambium and the cork cambium.
The wood is actually a secondary xylem. There are different types of wood on the basis of their composition ,and time of production.
Secondary Growth
Secondary Growth
Primary growth refers to the lengthening of roots and stems with the help of the apical meristem. Most dicotyledonous plants expand in girth in addition to main growth. This is referred to as secondary growth. The two lateral meristems, vascular cambium and cork cambium, are engaged in secondary growth.
A. Vascular Cambium:
Vascular cambium is the meristematic layer responsible for cutting off vascular tissues – xylem and phloem. It appears in patches as a single layer between the xylem and phloem in the young stem. It eventually forms a complete ring.
i) Formation of cambial ring:
The intrafascicular cambium is the cambium cells found between the primary xylem and primary phloem in dicot stems. The interfascicular cambium is formed when the cells of medullary rays next to this intrafascicular cambium become meristematic. As a result, a continuous ring of cambium forms.
ii) Activity of the cambial ring:
The cambial ring becomes active and starts cutting off new cells from both the inside and outside. Secondary xylem develops from cells cut off towards the pith, while secondary phloem develops from cells cut off towards the periphery. The inner side of the cambium is often more active than the outer side. As a result, secondary xylem is formed in greater quantities than the secondary phloem, forming a compact mass. The persistent development and accumulation of secondary xylem gradually crushes the primary and secondary phloems. The principal xylem, on the other hand, is mostly intact at or near the centre.The cambium sometimes produces a narrow band of parenchyma that runs in radial directions through the secondary xylem and secondary phloem. These are the secondary medullary rays.
iii) Springwood and autumn wood:
Cambium activity is influenced by a variety of physiological and environmental variables. Climate conditions in temperate zones are rarely consistent throughout the year. The cambium is particularly active in the spring and produces a significant number of xylary components with larger vessels. Springwood, also known as earlywood, is formed during this season. Autumn wood or latewood is formed when the cambium is less active in the winter and produces fewer xylary components with thin channels. Springwood is lighter in color and denser, whereas fall wood is darker and denser. An annual ring is made up of two types of wood that appear as alternate concentric rings.Annual rings in a cut stem can be used to estimate the age of a tree.
iv) Heartwood and sapwood:
The deposition of organic components such as tannins, resins, oils, gums, aromatic chemicals, and essential oils in the middle or innermost layers of the stem causes the majority of secondary xylem in ancient trees to be dark brown. These compounds make it tough, long-lasting, and resistant to bacteria and insects. Heartwood is a zone made up of dead components with heavily lignified walls. The stem is supported mechanically by the heartwood, which does not conduct water. Sapwood refers to the lighter-colored portion of the secondary xylem. It helps to transport water and minerals from the root to the leaf.
B.Cork Cambium:The outer cortical and epidermal layers break down as the stem grows in girth due to vascular cambium activity, and they must be replaced to create new protective cell layers. As a result, additional meristematic tissue is known as cork cambium or phellogen forms, usually in the cortical region, sooner or later. Phellogen is made up of several layers. It is made up of cells that are tiny, thin-walled, and practically rectangular. Both sides of the cell are cut off by Phellogen. Inner cells differentiate into secondary cortex or phelloderm, whereas exterior cells differentiate into cork or phellem. Due to suberin deposition in the cell wall, the cork is impenetrable to water. Secondary cortical cells are parenchymatous. The periderm is the combination of phellogen, phellem, and phelloderm.
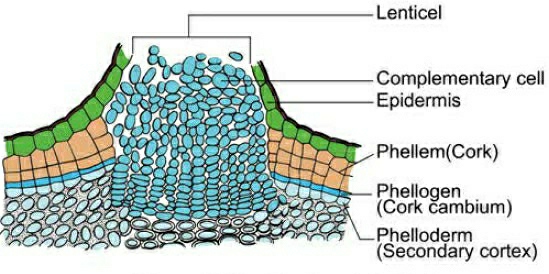
Pressure goes up on the remaining layers peripheral to phellogen due to cork cambium activity, and these layers eventually die and peel off. Secondary phloem is included in the phrase "bark," which is a non-technical term that refers to all tissues outside of the vascular cambium. Periderm and secondary phloem are two forms of tissue that makeup bark. Early or soft bark refers to bark that forms early in the season. Late or hard bark develops toward the end of the season. Instead of cork cells, the phellogen breaks off tightly packed parenchymatous cells on the outer surface in some areas. These parenchymatous cells quickly break the epidermis, generating lenticels, which are lens-shaped holes.Lenticels allow gas exchange between the outside atmosphere and the stem's internal tissue. Most woody trees have these.

C. Secondary Growth in Roots:
The vascular cambium in dicot roots is entirely secondary in origin. It develops from tissue right beneath the phloem bundles, a piece of pericycle tissue, and above the protoxylem, forming a complete and continuous wavy ring that eventually becomes circular. The next events are similar to those outlined before for a dicotyledon stem. Gymnosperm stems and roots also experience secondary growth. Monocotyledons, on the other hand, do not have secondary growth. Secondary growth usually results in a thickening of the root diameter due to the addition of vascular tissue. When cells in the residual procambium and sections of the pericycle begin to make periclinal divisions, secondary growth begins. Periclinal divisions begin only in the pericycle cells opposite the xylem sites. The vascular cambium is formed by the inner layer of cells. The pericycle is the outer layer that is retained. Around the primary xylem, the vascular cambium is continuous. The periclinal division of the vascular cambium continues. If the daughter cells divide towards the core of the root, they become secondary xylem cells, and if they divide towards the outer surface of the root, they become secondary phloem cells.The periderm, which comes from the pericycle and replaces the epidermis, forms an outer protective layer on certain roots. The pericycle restores its meristematic nature and divides periclinallyonce more and thus forms the phellogen or cork cambium. To the outside of the plant, the cork cambium generates phellum cells (cork cells). At maturity, these cells are lifeless. They've been suberized, which makes the cells water-resistant. Phellum cells in cross-section are neatly organized into files. The phelloderm, a mature tissue made up of live cells, is likewise produced by the cork cambium.