 ACME SMART PUBLICATION
ACME SMART PUBLICATION
Types of Movement
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
TYPES OF MOVEMENTS
Movements can be of 2 categories.
1.Non-Muscular Movements:
These movements persist in the animals in some of their cells.
(i) Protoplasmic Streaming: Streaming of protoplasm called cyclosis, has been seen in most of the cells such as leucocytes, Amoeba and other unicellular organisms.
(ii) Pseudopodial Movements: Leucocytes and macrophages move about in the tissues with the help of pseudopodia in the same manner as of Amoeba.
(iii) Flagellar Movements: The flagella of certain cells (e.g., choanocytes of poriferans) maintain by their ceaseless vibrations a regular current of water through the canal system of sponges. The flagella of certain gastrodermal cells circulate fluid in the coelenteron of Hydra by regular beating. Sperms swim in water or in female genital tract by flagellar movements.
(iv) Ciliary Movements: The cilia of the cells lining the trachea, oviducts and vasa efferentia propel dust particles, eggs and sperms in specific direction. The cilia of flame cells push waste material in excretory canals in flatworms.
The non-muscular ciliary locomotion is retained by some animal larvae such as the planula larva of coelenterates and the trochophore of annelids, and even some adults such as planarians.
2.Muscular Movements:
This depends upon the use of muscle fibres which have ability to exert force by alternate contraction and relaxation.
Most multicellular organisms have muscle fibres for moving different body parts or locomotion.
A muscle contraction does not always result in movement. It may at times maintain status quo, as in a fresh water mussel (mollusc), muscle contract to keep shell closed for safety.
Muscles
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Muscles
In human, muscular movements are involved.
Types:
There are three types of muscle tissue; striated or striped, nonstriated or unstriped or smooth, and cardiac, according to their location, structure and function.
Structure of Skeletal Muscle
The striated muscle forms 80% or more of the mass of soft tissues in a vertebrate body and found in the body wall and the limbs. It also occurs in the tongue, pharynx and beginning of oesophagus.
Skeletal muscle are having a connective tissue sheath on the outer side and is called epimysium.
A transverse section of it shows a number of bundles or fasciculi.
Each fasciculus is surrounded by connective tissue cover called perimysium.
Within a fasciculus, are present a large number of muscle fibres, each surrounded by connective tissue cover endomysium.
There is a broad band of fibrous connective tissue beneath the skin or around muscles called fascia.
Each muscle fibre is cylindrical, uniform in diameter. Sarcolemma is present on outer side and at places is invaginated to form T or transverse tubules. A skeletal or striated muscle fibre is multinucleated or syncytial.
Ultra Structure of Skeletal Muscle Fibre
Structure:
A striated muscle consists of long, narrow, cylindrical, unbranched fibres with blunt ends.
Each fibre is bounded by an elastic sarcolemma and contains many elongated, flattened nuclei characteristically located near the sarcolemma.
Multinucleate condition results from cell fusion. Hence, a striated muscle fibre is a syncytium.
The striated muscle fibres contain numerous mitochondria and glycogen granules for the supply of adequate energy.
The myofibrils of a striated muscle fibre show alternating dark and light cross bands, the striations, or stripes, hence the name of the muscle.
The dark bands are called anisotropic or A bands.
Each A band has at its middle a light zone termed Henson's line, or H zone.
The light bands are isotropic and are known as the isotropic or I bands.
Each I band is crossed through its centre by a dark membrane, the membrane of Krause, or Z line.
This membrane continues right across the whole fibre and joins the sarcolemma surrounding the fibre.
It seems to hold the myofibrils together and to carry the signals for the contraction of the fibrils inward from the T-tubules (transverse tubules).
The latter are invaginations of sarcolemma into the fibre adjacent to the Z lines.
The part of the myofibril between two successive Z lines functions as a contractile unit termed the sarcomere.
The sarcoplasm also contains a protein pigment myoglobin, which can take up, store, or give up oxygen like haemoglobin.
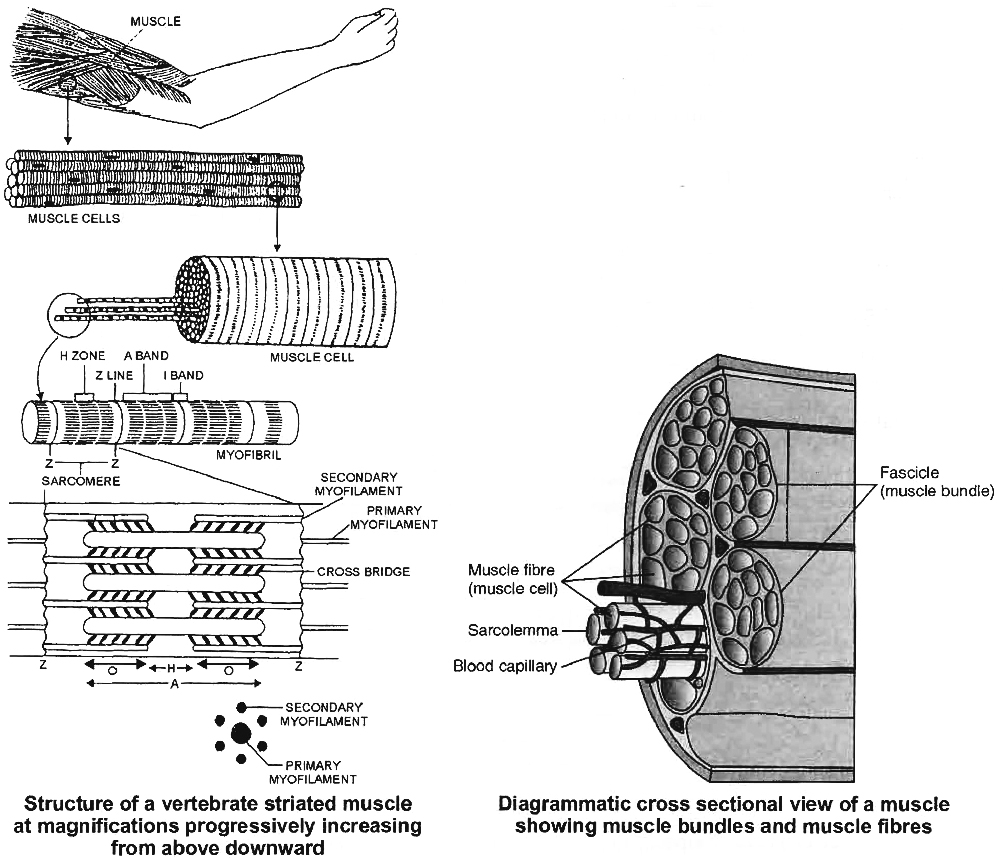
Electron microscope reveals that each sarcomere is a bundle of fine longitudinal myofilaments of two types : primary and secondary.
(i) Primary Myofilaments:
The primary myofilaments are thicker and confined to the A band only.
They are composed of the protein myosin; bear minute projections called cross-bridges of the protein meromyosin; and are free at both the ends.
(ii) Secondary Myofilaments:
The secondary myofilaments are thinner and occur in I bands, but extend for some distance into the A band between the primary myofilaments.
This partial overlapping of the primary myofilaments by the secondary myofilaments imparts dark appearance to the A bands.
The secondary myofilaments are composed of the proteins actin, tropomyosin and troponin; have a smooth surface; and are attached to the Z lines by one end, being free at the other end.
The secondary (actin) myofilaments are more numerous than the primary (myosin) myofilaments.
Six actin myofilaments surround each myosin myofilament and each actin myofilament is surrounded by three myosin myofilaments.
Mechanism of Muscle Contraction
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
MUSCLE CONTRACTION
When a nerve impulse (nerve action potential) reaches the synaptic end bulbs, it triggers exocytosis of synaptic vesicles.
In this process, the synaptic vesicles fuse with the plasma membrane and liberate ACh, which diffuses into the synaptic cleft between the motor neuron and the motor end plate.
When ACh binds to its receptor, a channel that passes small cations, most importantly Na+ opens.
The inrush of Na+ changes the resting membrane potential, which triggers a muscle action potential that travels along the muscle cell plasma membrane and initiates the events leading to muscle contraction.
Hanson and Huxley proposed that skeletal muscle shortens during contraction because thin filaments slide over thick filament.
Their model is known as the sliding filament mechanism of muscle contraction.
Sliding Filament Mechanism
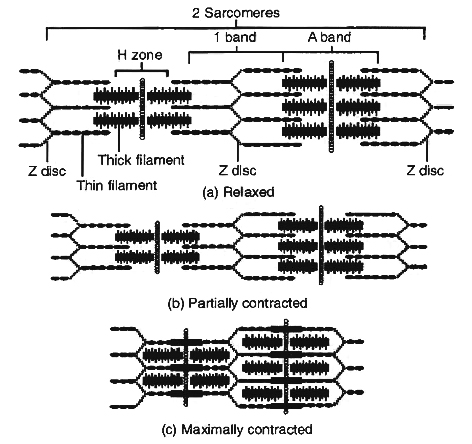
During muscle contraction, myosin heads pull on the thin filaments, causing them to slide inward towards the H zone at the centre of the sarcomere.
The myosin cross bridges may even pull the thin filaments of each sarcomere so far inward that their ends overlap in the centre of the sarcomere.
As the thin filaments slide inward, the Z discs come towards each other, and the sarcomere shortens, but the lengths of the thick and thin filaments do not change.
The sliding of the filaments and shortening of the sarcomeres cause shortening of the whole muscle fibre and ultimately the entire muscle.
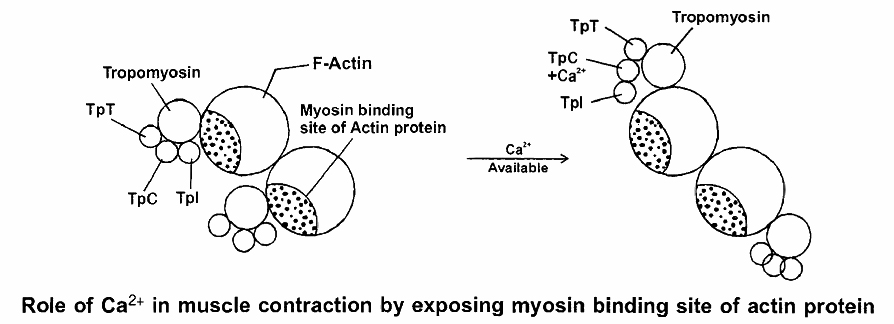
Role of Calcium and Regulator Proteins
An increase in Ca2+ concentration in the sarcoplasm starts filament sliding, while a decrease turns off the sliding process.
When a muscle fibre is relaxed (not contracting), the concentration of Ca2+ in its sarcoplasm is low.
This is because the sarcoplasmic reticulum (SR) membrane contains Ca2+ active transport pumps that move Ca2+ from the sarcoplasm into the SR.
Ca2+ is stored or sequestered inside the SR.
As a muscle action potential travels along the sarcolemma and into the transverse tubule system, however, Ca2+ release channels open in the SR membrane.
As a result, Ca2+ floods into the sarcoplasm around the thick and thin filaments.
The Ca2+ released from the sarcoplasmic reticulum combine with troponin, causing it to change shape.
This shape change moves the troponin -tropomyosin complex away from the myosin-binding sites on actin.
Troponin has 3 units (tri units structure)
1. TpT - tropomyosin binding troponin
2. TpC - calcium binding protein
3. Tpl - Inhibitor i.e., blocks myosin binding site of actin proteins
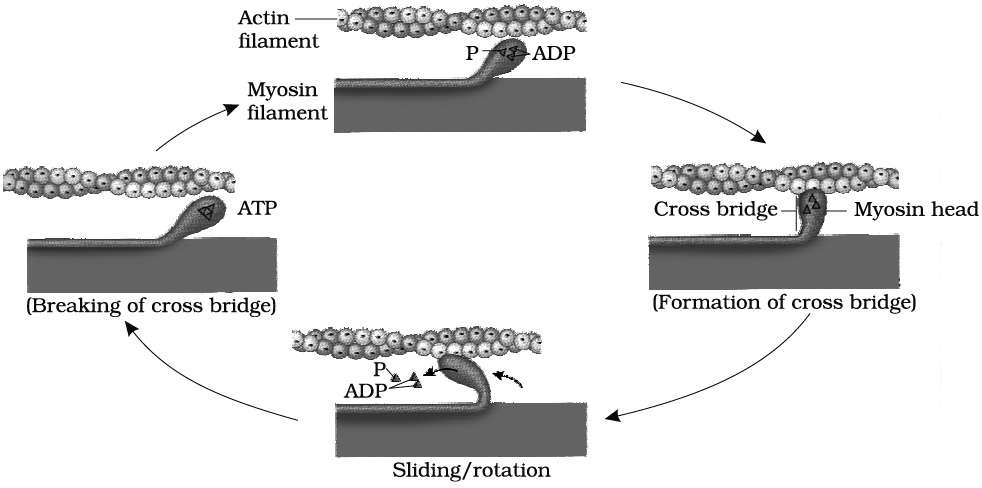
The Power Stroke and the Role of ATP
As we have seen, muscle contraction requires Ca2+ ions and energy in the form of ATP. The sequence of events during sliding of the filaments are
(i) While the muscle is relaxed, ATP attaches to ATP-binding sites on the myosin cross bridges (heads). A portion of each myosin head acts as an ATPase, an enzyme that splits the ATP into ADP + P (phosphate group) through a hydrolysis reaction. This reaction transfers energy from ATP to the myosin head, even before contraction begins. The myosin cross bridges are thus in an activated (energized) state.
(ii) When the sarcoplasmic reticulum releases Ca2+, its level rises in the sarcoplasm. Rise in Ca+2 binds with troponin and change its configuration that moves away tropomyosin from its blocking position.
(iii) The activated myosin heads spontaneously bind to the myosin-binding sites on actin.
(iv) The shape change that occurs as myosin heads bind to actin produces the power stroke of contraction. During the power stroke, the myosin heads swivel toward the centre of the sarcomere, like the oars of a boat during rowing. This action draws the thin filaments past the thick filaments toward the H zone. As the myosin heads swivel, they release ADP.
(v) Once the power stroke is complete, ATP again combines with the ATP-binding sites on the myosin heads. As ATP binds, the myosin head detaches from actin.
(vi) Again, the myosin ATPase splits ATP, transferring its energy to the myosin ATPase splits head, which returns to its original upright position .
(vii) The myosin head is then ready to combine with another myosin-binding site further along the thin filament.
The steps (iii) through (vii) repeats over and over as long as ATP is available and the Ca2+ level near the thin filament is high.
The myosin heads keep rotating back and forth with each power stroke, pulling the thin filaments toward the H zone.
At anyone instant, about half of the myosin heads are bound to actin and are swiveling.
The other half are detached and preparing to swivel again.
This continual movement of myosin heads applies the force that draws the Z discs toward each other, and the sarcomere shortens.
The myofibrils thus contract and the whole muscle fibre shortens.
During a maximal muscle contraction, the distance between Z discs can decrease to half the resting length.
H-line and M-line disappear, I-band almost disappears, A-band remains constant, but the power stroke does not always result in shortening of the muscle fibres and the whole muscle.
Contraction without shortening is called an isometric contraction, for example, in trying to lift a very heavy object.
The myosin heads (cross bridges) swivel and generate force, but the thin filaments do not slide inward.
Relaxation: Two changes permit a muscle fibre to relax after it has contracted.
First, acetylcholine is rapidly broken down by an enzyme called acetylcholinesterase (AChE).
When action potentials cease in the motor neuron, release of ACh stops, and AChE rapidly breaks down the ACh already present in the synaptic cleft.
This ends the generation of muscle action potentials, and the Ca2+ release channels in the sarcoplasmic reticulum membrane close.
Second, Ca2+ active transport pumps rapidly remove Ca2+ from the sarcoplasm into the sarcoplasmic reticulum, where molecules of a calcium-binding protein, appropriately called calsequestrin, bind to the Ca2+.
With this, the tropomyosin-troponin complex move back over the myosin binding site of actin which prevents further binding of myosin head to actin and the thin filaments slide back to their normal relaxed position.
All or None Principle
A minimal strength of a stimulus required to cause the contraction of a muscle fibre brings about maximum contraction, and no further increase in contraction would occur by increasing the strength of the stimulus.
Single Muscle Twitch
A Single, quick isolated contraction of a muscle fibre to a single stimulus of threshold value is called single muscle twitch in the laboratory experiments.
Energy Source of Muscle Contraction
Energy for muscle contraction is provided by hydrolysis of ATP by myosin ATP-ase enzyme.
This hydrolysis produces ADP, inorganic phosphate and energy (used in muscle contraction).
Phosphocreatine donates its high energy and phosphate to ADP, producing ATP.
Phosphocreatine serves as an energy source for a few seconds for metabolic processes in the muscle cells to begin to produce greater quantities of ATP.
Phosphocreatine is again formed in relaxing muscle by using ATP produced by carbohydrate oxidation.
ATP + H2O ![]() ADP + Pi + Energy
ADP + Pi + Energy
(Inorganic phosphate)
Phosphocreatine + ADP ![]() ATP + Creatine
ATP + Creatine
At the end of muscle contraction, the conversion of ADP into ATP takes place.
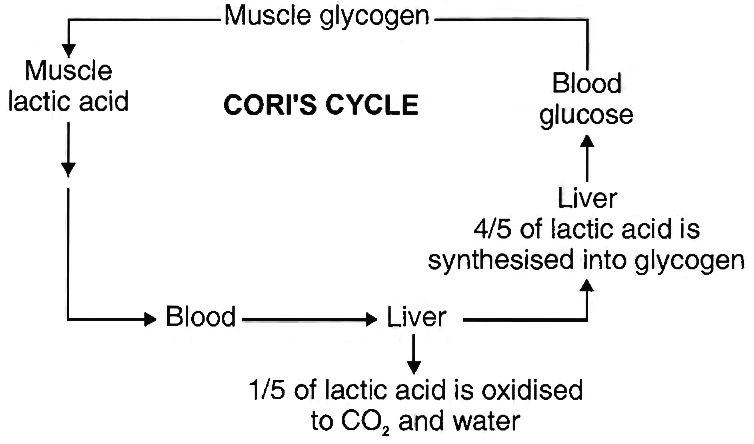
The muscle is rich in glycogen which is broken down into lactic acid through a series of reactions (glycolysis) and liberates energy.
Some of this energy is used for the reformation of phosphocreatine and also for the conversion of 4/5th of lactic acid back into glycogen.
The 1/5th of lactic acid is oxidised to water and carbon dioxide.
These reactions taking place in the muscle and liver, are proposed by Cori and Cori, hence known as Cori's cycle.
Rigor Mortis
Extreme rigidity of body after death is called rigor mortis. It is due to complete depletion of ATP and phosphocreatine.
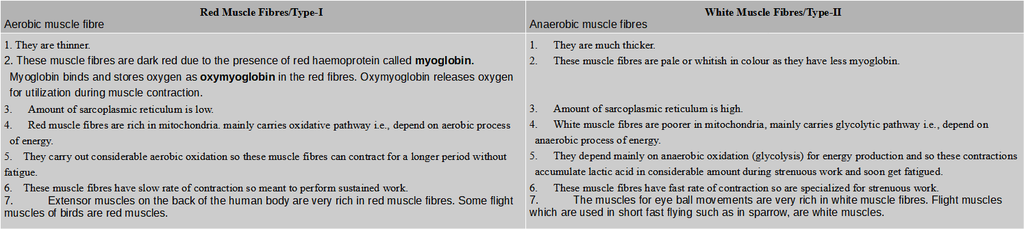
Red and White Muscle Fibres
Birds and mammals have two kinds of striated muscle fibres in their skeletal muscles; red or slow muscle fibres and white or fast muscle fibres.
Differences between red muscle fibres and white muscle fibres
Contraction in Smooth Muscles:
In comparison with contraction in a skeletal muscle fibre, contraction in a smooth muscle fibre starts more slowly and lasts much longer Troponin is absent in smooth muscle so they have regulator protein called Calmodulin that binds to Ca2+ in the cytosol.
Using ATP, myosin head part can bind to actin & contraction can occur.
Most smooth muscle fibres contract or relax in response to action potentials from the autonomic nervous system.
Contraction in Cardiac Muscles:
Cardiac muscle fibres have the same arrangement of actin & myosin and the same bands, zones and Z discs as skeletal muscle fibres.
Gap junctions allow muscle action potential to spread from one muscle fibre to another.
As a consequence, when a single muscle fibre is stimulated, all the other fibres in the network become stimulated as well.
Thus, each network contracts as a functional unit.
Cardiac muscle tissue has a long refractory period and can use lactic acid produced by skeletal muscle fibres to make ATP.
Skeletal System
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
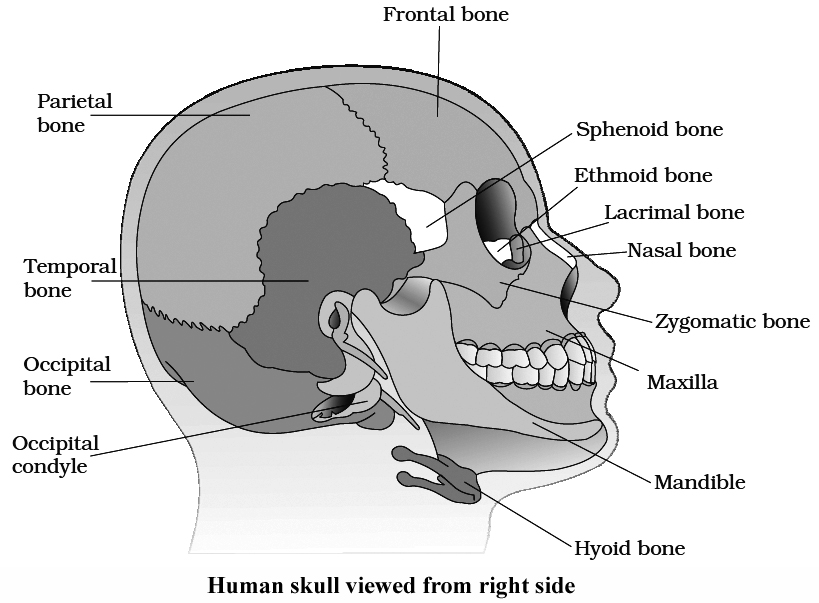
SKELETAL SYSTEM IN HUMAN
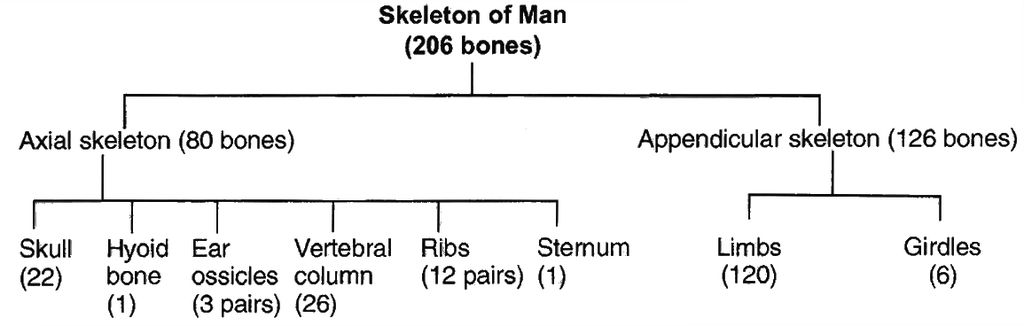
The skeletal system is divided into two main parts.
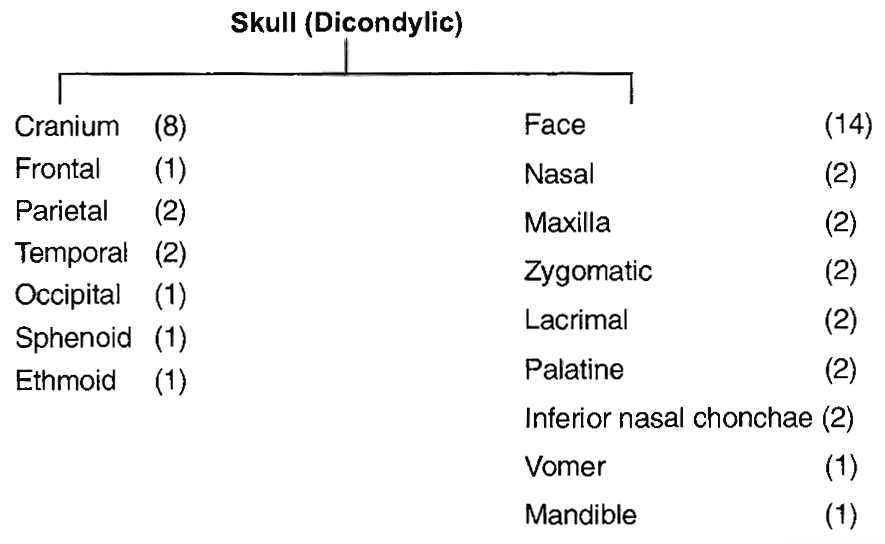
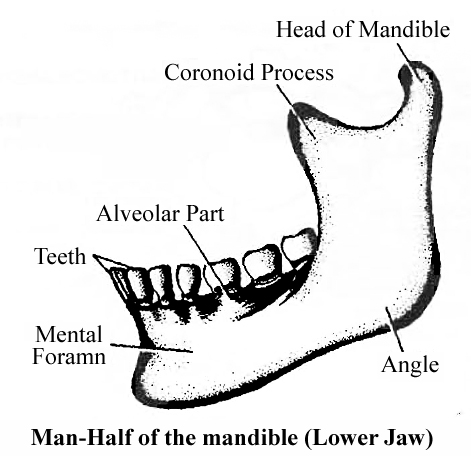
(i) Axial Skeleton. It lies along the principal axis of the body. It includes skull, vertebral column, ribs, sternum and ribs.
(ii) Appendicular Skeleton. It is made up of the girdles and limb bones.


Hyoid
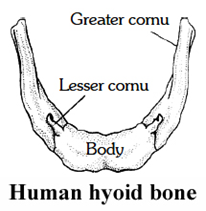
It serves as a point of attachment for some of the muscles of the tongue and floor of the mouth but does not articulate with any other bone.
Ear Ossicle
Ear ossicles are three in number

VERTEBRAE
Vertebral column protects the spinal cord, supports the head and serves as the point of attachment for the ribs and musculature of the back.
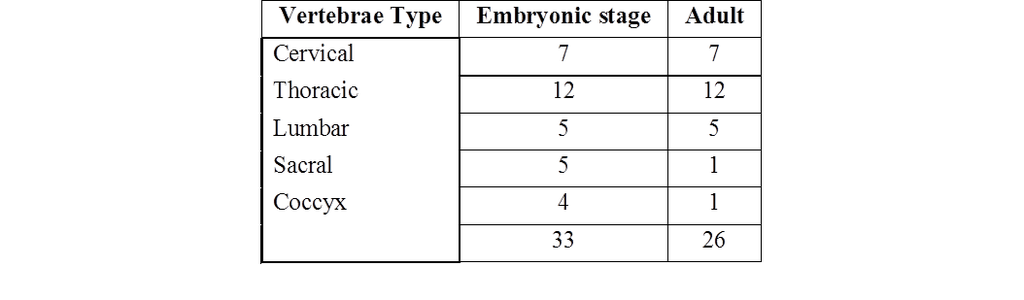
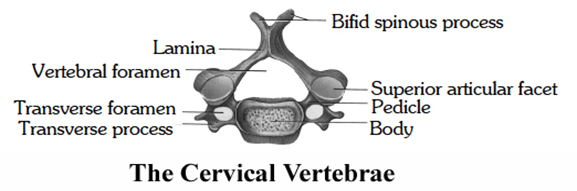
Cervical vertebrae
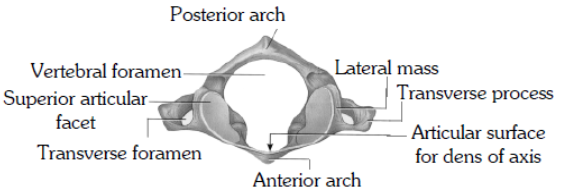
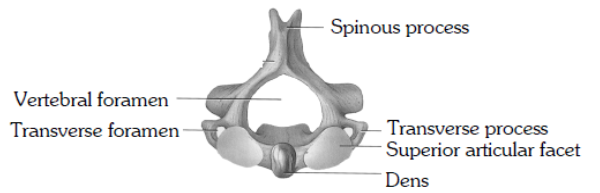
(i) 1, 2, 7 cervical vertebrae are atypical and 3 to 6 are typical.
(ii) The transverse processes are perforated by foramina (foramen transversarium) for the passage of the vertebral arteries except in VII Vertebrae.
(iii) The first cervical vertebra or atlas supports the head and consists of a complete ring of bone. On its upper surface it presents kidney-shaped facets for articulation with the condyles of the occipital bone, forming a condyloid joint, the atlanto-occipital joint, at which the nodding movements of the head take place. The atlas articulates with the second cervical vertebra.
(iv) The second cervical vertebra or axis is the pivot on which the atlas turns in the rotary movements of the head. From the body of the axis a process of bone rises which is called the odontoid peg which articulates with the back of the anterior arch of the atlas and is held in position by the transverse ligament of the atlas. The lateral masses of the atlas articulate with corresponding facets on the axis placed on each side of the odontoid peg. The atlas moves round the odontoid peg of the axis, forming a pivot joint at which the head rotates.
(v) The seventh cervical vertebra is the first vertebra with an undivided spinous process. This process has a tubercle at its tip. It forms a distinct projection in the neck and can be seen at the lower part of the back of the neck. Because of this characteristic the bone is called the vertebra prominens.
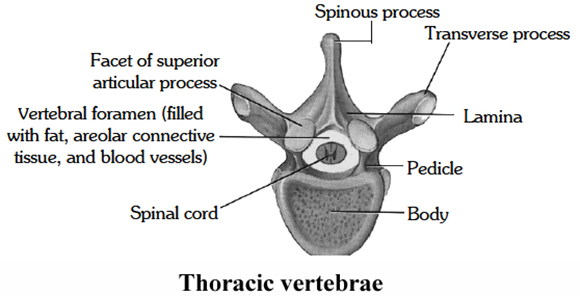
Thoracic Vertebrae
(i) Thoracic 2-8 are typical 1, 9, 10, 11, 12 are atypical.
(ii) These are larger than the cervical and they increase in size as they extend downwards.
(iii) The body is heart-shaped, with facets on each side for attachment of the ribs.
(iv) The neural arch is relatively small, the spinous process is long and is directed downwards.
(v) The transverse processes which help to support the ribs are thick, strong and carry articular facets for the ribs.
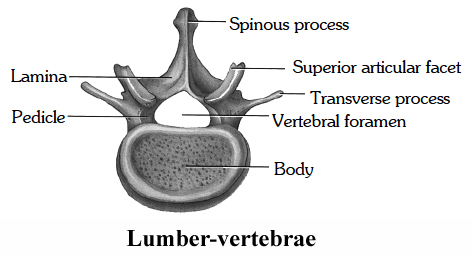
Lumbar Vertebrae
(i) These are the largest vertebrae as compared with the bodies of the other vertebrae and is kidney-shaped.
(ii) The spinous process is broad and hatchet-shaped.
(iii) The transverse processes are long and slender.
(iv) The fifth lumbar vertebra articulates with the sacrum at the lumbo-sacral joint.
Sacrum
(i) It is a triangular bone situated at the lower part of the vertebral column, wedged in between the two innominate bones and forming the back of the pelvic cavity.
(ii) The base of the sacrum lies above and articulates with the fifth lumbar vertebra, forming a typical intervertebral joint.
(iii) The junction between the fifth lumbar vertebra and the sacrum forms the sacro-vertebral angle.
(iv) At the extremities of transverse ridges, on each side, sacral foramina are present for the passage of nerves. The apex of the sacrum articulates with the coccyx. At the sides the sacrum articulates with the innominate bones, forming the right and left sacro-iliac joint.
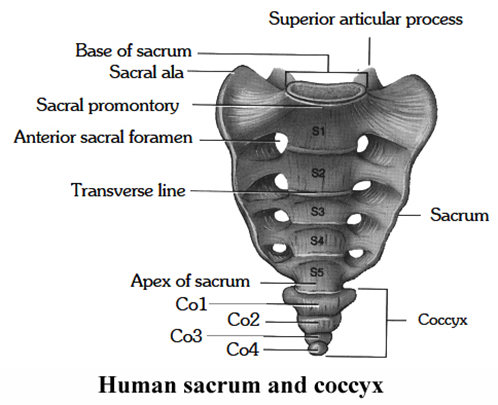
Coccyx
It is composed of four or five rudimentary vertebrae, fused to form one bone. It articulates above with the sacrum.
Joints
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
THE JOINTS
A bone joint or articulation may be defined as the junction of two bones. The study of such joints is known as arthrology.
There are three principal types of bone joints.
(i) Fibrous joints or immovable or fixed joints/synarthrosis
(ii) Cartilaginous joints/Amphiarthrosis
(iii) Synovial joints/Diarthrosis
(i) Fibrous Joints.
These joints are immovable or fixed.
They do not show any movement due to the presence of strong and tough white collagenous fibres and there is no joint cavity.
These joints include:
(a) Sutures: Found between skull bones, articulating bones are held together by white fibrous tissue.
(b) Gomphoses : Teeth in mandibles and in maxillary bones.
(c) Shindylases : One bone fits into slit in another e.g., ethmoid bone in vomer.
(ii) Cartilaginous Joints.
They are slightly movable joints.
Discs of white fibrocartilage, strong but more elastic and compressible than the white fibrous tissue, hold the bones together at the joints between the bodies of the vertebrae, at the symphysis pubis, and between the sternum and ribs.
The bones make some movements at such joints through compression of the discs of the cartilage.
(iii)Synovial Joints.
Synovial joints are of different types depending upon the nature of articulation and degree of movement.
Synovial joints are of the following types:
(a) Ball and socket joints ( = Enarthroses) : The 'head' of one bone fitting with the 'socket' of the other bone and allowing free movement in all planes; e.g. shoulder joint and hip joint.
(b) Hinge joints ( = Ginglymi) : The perfect joints which allow the movements only in a single plane. e.g. elbow joint, knee joint and ankle joint
(c) Pivotal joints (=Rotary joints or rotaria) : One of the two bones is fixed in its place and bears a peg like process over which rotates the other bone; e.g. atlas along with the skull rotating over the odontoid process of axis vertebra in mammals.
(d) Saddle joints : It is similar to ball and socket joints but are poorly developed and movements are comparatively less free, e.g. the joint between the metacarpal of thumb with the carpals below.
(e) Gliding joints: The joints which permit sliding of the articulating bones on each other; e.g. joint between the zygapophyses of successive vertebrae, and between sternum and clavicle.
(f) Angular joints (= or ellipsoid or condyloid) : These joints allow the movements in two directions, i.e., side to side and back and forth; e.g., metacarpophalangeal joints.
Disorder of Muscular and Skeletal System
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Disorder of Muscular and Skeletal System
1.The rheumatoid arthritis:
It is diagnosed by the presence of rheumatoid factor (a type of immunoglobulin IgM).
It is the primary symptom of inflammation of synovial membrane.
If it is left untreated, then the membrane thickens and synovial fluid increases, exerting pressure that causes pain.
The membrane then starts secreting abnormal granules, called pannus, which after accumulating on the surface of the cartilage, causes its erosion.
As a result, the fibrous tissues are attached with the bones and become ossified, making the joints immovable heat treatment and physiotherapy pain and inflammation and in extreme cases, replacement of the damaged joints is recommended
2. Osteoarthritis:
It is a degenerative joint disease characterised by the degeneration of the articular cartrlage and proliferation of new bones.
Usually, affected joints are of spine, knees and hands.
3. Gouty Arthritis or Gout:
It is caused either due to excessive formation of uric acid, or inability to excrete it.
It gets deposited in joints as monosodium salt.
4. Osteomalacia and Rickets:
Osteomalacia, called rickets when it occurs in childhood.
In this disease disorder this bones contain insufficient amounts of calcium and phosphorus.
The cause of kidney disease, vitamin D deficiency, and an inherited defect.
5. Osteoporosis:
Osteoporosis is a disease in which bone loses minerals and fibres from its matrix.
Individuals taking hydrocortisone for arthritis, allergies, or other disorders are especially prone to bone loss.
6. Bursitis:
The bursae of joints often become inflamed, a condition known as bursitis.
The inflammation can be caused by a physical injury or by constant pressure to the same joint over a long period of time.
7. Dislocation:
A dislocation is a displacement of the articular surfaces of a joint; it usually involves a damage to the ligaments surrounding the joint.
Most dislocations result from falls, blows, or extreme exertion and are most often seen in the joints of the thumb, fingers, knee or shoulder.
Symptoms of dislocation include swelling, pain, and loss of motion.
8. Sprain and Strains:
A sprain is a twisting of a joint without dislocating it.
Such an injury causes damage to ligaments and also often damages tendons, muscles, blood vessels, and nerves.
Severe sprains are quite painful and require immobilization during the healing process.
In contrast to a sprain, a strain is a less severe stretching or twisting of a joint.
Muscles and tendons may be stretched and become somewhat painful, but only minor damage is done to the tissues of the joint.
9. Myasthenia gravis:
Auto immune disorder affecting neuromuscular junction leading to fatigue, weakening and paralysis of skeletal muscle.
10. Muscular dystropy:
Progressive degeneration of skeletal muscle mostly due to genetic disorder.
11. Tetany:
Rapid spasm (wild contraction) in muscle due to low Ca++ in body fluid.
