 ACME SMART PUBLICATION
ACME SMART PUBLICATION
cyclic and non cyclic photophosphorylation and The Electron Transport
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Cyclic and Non cyclic photophosphorylation
Cyclic photophosphorylation :
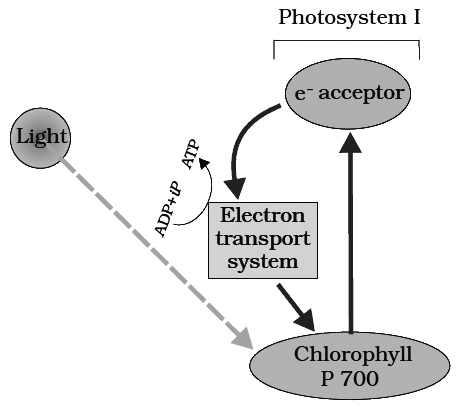
The process of cyclic photophosphorylation involves only photosystem I.
A photon of light excites an electron of chlorophyll a molecule (of PS I) and the excitation energy is eventually passed on to the reaction centre P700.
This excited electron is expelled from the chlorophyll molecule and is accepted by an electron acceptor Fe-S.
By initiating this electron transfer chain, P700 molecule becomes oxidised.
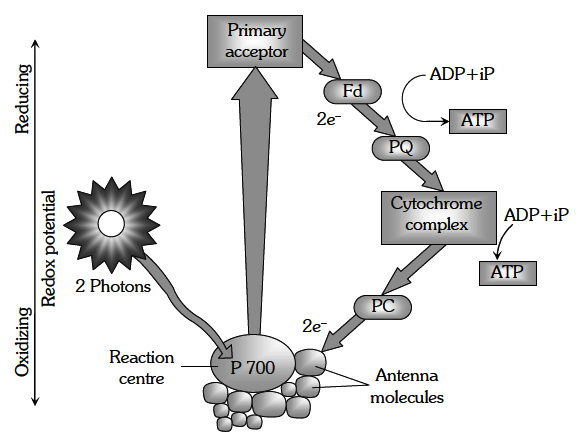
Fe-S passes this electron to other acceptors in the following sequences : Ferredoxin, Plastoquinone, Cytochrome b6-f complex and plastocyanin.
The oxidised P700 serves as a natural electron acceptor from plastocyanin. All the above electron acceptors are in oxidised state before accepting electrons (therefore these act as electron acceptors) and get reduced after accepting the electron.
As the electron is transferred from one acceptor to another, certain amount of energy is released. This energy is used to generate A TP (while passing between Fd and PQ and/or PQ and cyt. b6-f complex).
When only PSI is functional, the electron is circulated within the photosystem and the phosphorylation occurs due to cyclic flow of electrons.
A possible location on where it occurs is in the stroma lamellae.
The stroma lamellae membrane lacks PSII as well as NADP reductase enzyme.
The excited electron does not pass on to NADP+ but is cycled back to the PSI complex through electron transport chain.
The cyclic flow hence, results only in the synthesis of ATP, but not of NADPH + H+.
Cyclic photo phosphorylation also occurs when only light of wavelengths beyond 680 nm are available for excitation.
(b) Non-cyclic photophosphorylation:
It involves photosystem I as well as photosystem II. Both these systems absorb light photons. For convenience, the non-cyclic photophosphorylation is described here in following steps:
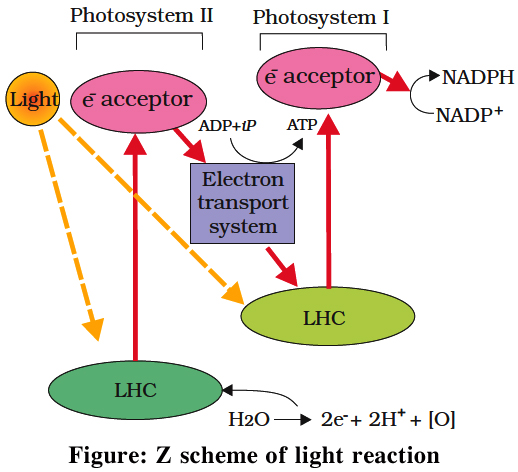
(i) When P680 absorbs a photon of red light, it gives out an electron, and thus becomes positively charged. This electron from P680 passes to P700 through a series of electron carriers, like Phaeophytin (Primary acceptor), plastoquinone, cytochrome b6-f complex, plastocyanin.
This movement of electron is downhill in terms of redox potential scale.
In this series, enough energy is released when the electron is transferred from PQ to cytochrome b6-f complex. It is utilized to synthesize ATP.
(ii) Simultaneously, electron in the reaction centre of PS I also get excited when they receive red light of 700 nm and these electrons are transferred to another receptor having high redox potential. These electrons then move downhill again, this time to a molecule of NADP+. This addition of electron and a proton, reduce it to NADPH + H+.
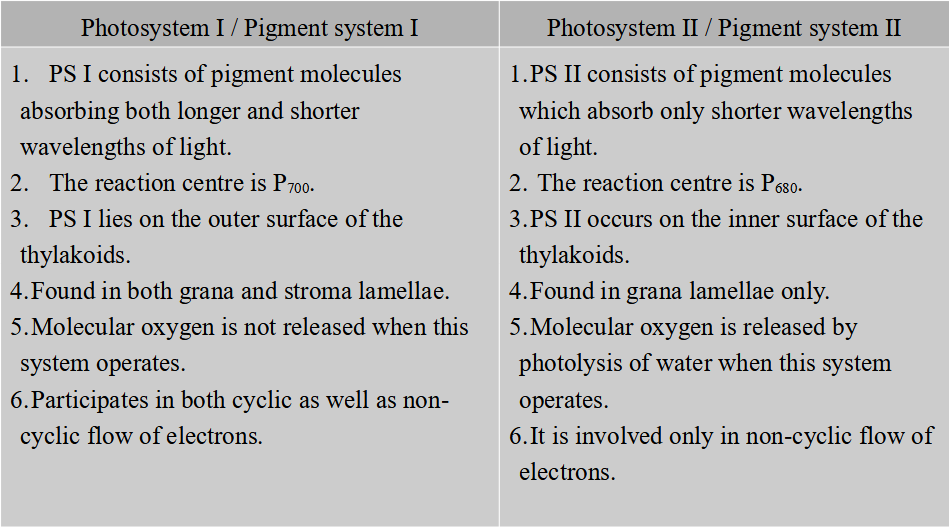
Differences between Cyclic and Non-cyclic Photophosphorylation

Prior knowledge of the photosynthesis and early experiments
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Prior knowledge of the photosynthesis and early experiments
Some important events related to photosynthetic process are described below.
(1) Stephan Hales was the first to prove that gaseous constituents of the air and light also contribute to the building up of the plant body.
(2) Joseph Priestley concluded that plants purify the impure air released by animals. Joseph Priestley (1733-1804) in 1770 performed a series of experiments that revealed the essential role of air in the growth of green plants. Priestley discovered oxygen in 1774.
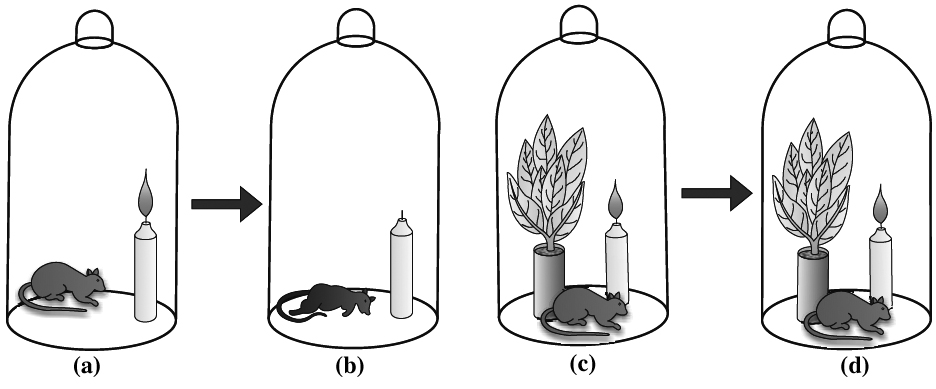
Priestley observed that a candle burning in a closed space -a bell jar, soon gets extinguished (Figure a, b, c, d). Similarly, a mouse would soon suffocate in a closed space. He concluded that a burning candle or an animal that breathe the air, both somehow, damage the air. But when he placed a mint plant in the small bell jar, he found that the mouse stayed alive and the candle continued to burn. Priestley hypothesised as follows: Plants restore to the air whatever breathing animals and burnning candles remove.
(3) Jan Ingenhousz mentioned, that in sunlight green leaves give out dephlogisticated air (air rich in oxygen) and in dark these green parts give out phlogisticated air (air rich in CO2) and make the air impure.
(4) Jean Senebier discovered that green plants utilize carbon dioxide. It was thus a great land mark in the discovery of photosynthesis.
(5) N.T. de Saussure proved that the volume of carbon dioxide consumed is equal to the volume of oxygen liberated. He also showed that water is essential requirement in photosynthesis.
(6) Pelletier and Caventou named the green coloured substance in the leaves as chlorophyll.
(7) F.F. Blackman observed that the process of photosynthesis has two steps : a photochemical reaction which proceeds only in the presence of light and a dark reaction for which light is not necessary. Besides this, he also proposed the law of limiting factor.
(8) Warburg performed Flashing experiment on Chlorella.
(9) Emerson and Arnold, with the help of experiments determined that the light reaction of photosynthesis has two distinct photochemical processes.
(10) Robert Hill's experiment on Stellaria media demonstrated that in the presence of sunlight, water and a suitable hydrogen acceptor, isolated chloroplasts release oxygen, even if carbon dioxide is absent. This experiment is considered to be equivalent to light reaction. Hill's oxidants are hydrogen acceptors. The common ones are ferricyanide, benzoquinone and dichlorophenol indophenol(DCPIP), while NADP+ is natural H+ acceptor in photosynthesis.
(11) Van Niel, based on his studies of purple and green sulphur bacteria demonstrated that hydrogen released from a suitable oxidisable compounds reduces CO2 to carbohydrates and proposed that water is the source of O2.
(12) Ruben, Hassid and Kamen using radioactive oxygen (O18) confirmed that the source of oxygen in photosynthesis is water.
(13) Julius von Sachs (1854) found that the green parts in plants is where glucose is made and glucose is usually stored as starch.
(14) T.W. Engelmann experimented on Cladophora and Spirogyra. He used a prism to split light into its components and illuminated the green alga placed in a suspension of aerobic bacteria. He observed the accumulation of bacteria in the region of blue and red light of the split spectrum. Thus, he described the first action spectrum of photosynthesis.
(15) Melvin Calvin and his coworkers discovered various reactions involved in the conversion of carbon dioxide into carbohydrates using C14O2. These reactions are collectively known as C3 cycle or Calvin cycle. He was awarded Nobel Prize for this work in 1961.
(16) M.D. Hatch and C.R. Slack discovered a new series of reactions involved in photosynthetic CO2 assimilation in many tropical plants. In these plants the first stable compound in photosynthetic process is a 4-carbon compound, hence, this series of reactions is known as C4 cycle.
(17) Hill and Bendall proposed the scheme of light reaction.
(18) Huber et. al. studied three dimensional structure of reaction center of bacteria Rhodopseudomonas viridis and got Nobel prize.
where does photosynthesis occur?
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Photosynthesis occur
In higher plants chloroplast is semiautonomous, endosymbiont with three membrane system. It is made up of three main parts: Envelope, Matrix and Thylakoid.
Envelope: It is made up of outer membrane ±porins, inner membrane, in between outer and inner membrane is the periplastidal space.
Semi fluid matrix known as stroma, which contain double stranded circular DNA, 70S ribosome,
Carboxylation enzyme RUBISCO, and PEP carboxylase enzyme Mg, Cl, Mn etc.
Thylakoid is membrane lined flattened sac like structure unite to form Grana.
Each thylakoid in grana is known as grana thylakoid, inside it contains the lumen for photolysis of water with the help of Mg, Mn, Cl, Ca and number of electron carriers.
Each grana interconnected with stroma lamellae. Inside it contains space known as fret channel.
Thylakoid membrane possesses 200 to 400 pigment molecule associated with certain protein known as CAB together known as quantosomes or light harvesting complex.
Thylakoid membrane also possess F0 -F1 particle or coupling factor and number of electron carriers arranged based on their redox potential
Photosynthetic Pigments
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Photosynthetic Pigments
There are mainly three types of pigments: chlorophylls, carotenoids and phycobilins (in BGA and red algae).
(1) Chlorophylls (green pigments)
The chlorophylls are main pigments which plants use in Photosynthesis.
They are specialized lipid molecules embedded in thylakoid membrane.
They are the main pigment concerned with harvesting or trapping of solar energy. Arnof and Allen (1966) recognised nine types of chlorophyll – (i) chlorophyll a; (ii) chlorqphyll b, (iii) chlorophyll c, (iv) chlorophyll d; (v) chlorophyll e; (vi) bacteriochlorophyll a, (vii) bacterio -chlorophyll b, (viii) chlorobium chlorophyll 650 and (ix) chlorobium chlorophyll 666.
Out of these, the two important types of chlorophyll found in green plants are chlorophyll a and
Of these, chlorophyll a is the primary photosynthetic pigment.
It is found in all photosynthetic eukaryotes and blue green algae (cyanobacteria).Light energy absorbed by all other types of chlorophylls and accessory pigments is ultimately transferred to chlorophyll a.
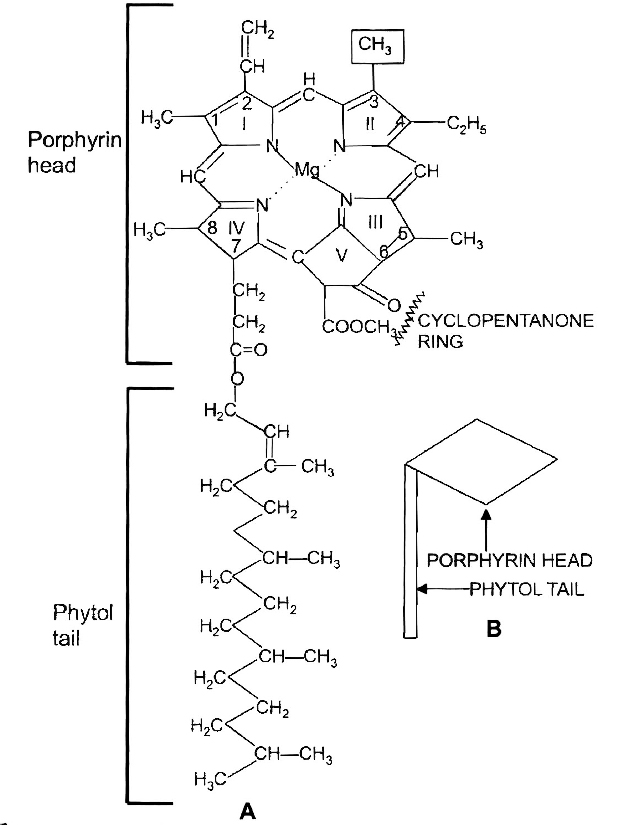
(a) Chlorophyll a : The chlorophyll a has a empirical formula as C55H72O5N4 Mg. The molecule is distinguishable into a 'head' of size 15 × 15 Å and a 'tail' of 20 Å length. The 'head' is made up at porphyrin, (a tetrapyrrole closed ring derivative) and the 'tail' of phytol (C20H39OH).
There is a 5th isocyclic ring of cyclopentanone. A non-ionic magnesium atom is held within tetrapyrrole ring by two covalent and two co-ordinate bonds. There is vinyl group at carbon-2 position and methyl at carbon-3. The chlorophyll a absorbs violet blue and red wavelengths of the spectrum giving peaks at 430 and 662 nm respectively. It is found in all photosynthetic organisms except bacteria. Protoclorophyll is its precursor.
(b) Chlorophyll b : Its empirical formula is C55H70O6N4 Mg. It is similar to chlorophyll a except in having a formyl (CHO) group instead of methyl (CH3) at carbon-3 position of the tetrapyrrole ring. It absorbs blue and orange wavelengths giving peaks at 430 and 644 nm.
(c) Chlorophyll c: It lacks phytol esterification. It is found in brown algae, diatoms and dinoflagellates.
(d) Chlorophyll d: It has formyl group at carbon-2 position. It is reported from red algae.
(e) Chlorophyll e: It has been reported from Xanthophyceae members like Vaucheria.
(f) Bacteriochlorophyll: The molecular formula of bacteriochlorophyll-a is C55H74O6N4 Mg. Its structure is similar to chlorophyll a except presence of acetyl group instead of vinyl at carbon-2 position.
(g) Chlorobium chlorophyll: It has hydroxy-methyl group (CH3CHOH) at carbon-2 position in the tetrapyrrole nucleus.
(2) Carotenoid pigments (Accessory or shield pigments):
Carotenoids are yellow to orange lipid compounds which occur in almost all higher plants. Carotenoids are of two types-carotenes and xanthophylls.
(a) Carotenes are reduced molecules (C40 H56). They are of several types like a and b.
b-carotene is the most widespread and important carotene associated with chlorophyll in photosynthetic organisms. It is orange-yellow in colour. Vertebrates are able to break each molecule of b-carotene in two molecules of vitamin A during digestion. Red colour of tomatoes is due to a carotene called lycopene.
(b) Xanthophylls. These pigments, besides carbon and hydrogen also contain oxygen, e.g., lutein (C40H56O2), cryptoxanthin (C40H56O), etc.
Carotenoids have at least two major roles in photosynthesis: (i) they transfer the light energy absorbed by them to chlorophyll molecules, hence are called antenna molecules. They absorb light in blue violet range and (ii) they protect plant from excessive heat and prevents photooxidation (oxidative destruction by light) of chlorophyll.
(3) Phycobilins (Biliprotein):
Phycobilins are another group of accessory pigments found in red and blue-green algae. They have open chain tetrapyrrole structure and are hot water-soluble pigments. Phycobillins can be classified into two types: (i) Phycoerythrins, (ii) Phycocyanins.
Usually both these types occur together, but their proportion may vary according to species and environment.
Mechanism of Photosynthesis
The two major steps of photosynthesis are -Light or Hill reaction and dark or Blackman reaction.
Concept Builder
Flashing (Intermittent) light experiment was performed by Warburg. He established that the rate of photosynthesis was found to be greater in intermittent light as compared to continuous light. It indicates the existence of two steps in photosynthesis, a light dependent reaction and another light independent reaction (the dark reaction, enzymatic reaction).
Photosynthetic Units / Pigment Systems
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Photosynthetic Units
These are group of pigment molecules which takes part in change of light energy into chemical energy.
Each photosynthetic unit has a reaction center (photocentre) of specific chlorophyll a molecule like P700 and P680, which absorb light energy of long wavelength.
These can release electron upon absorption of energy.
Reaction center is surrounded by a number of light harvesting pigment molecules.
These are those photosynthetic pigments which absorb photons of different wavelength.
Harvesting molecules occur in the form of specific complexes called LHC I and LHC II.
Harvesting molecules are of two types :
I. Antenna molecule : Those occurs on outer side of photosynthetic unit. They absorb photons of different wavelength depending upon their absorbance maxima. They get excited and energy is transferred to core molecule through resonance transfer.
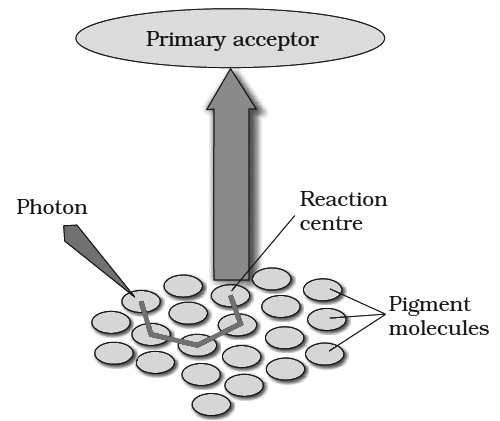
II. Core molecules: They are pigment molecules which are present around the reaction center. They transfer the energy to reaction center, so the reaction center gets excited.
(3) Evolution of oxygen :
Equations of photosynthesis proposed before 1930s gave the impression that the source of oxygen was carbon dioxide.
However, experiments of C.B. Van Niel performed during 1930s showed that oxygen gas liberated during photosynthesis is released as a result of photolysis of water.
While comparing photosynthetic activity of sulphur bacteria and higher plants, Van Niel found that sulphur bacteria assimilate carbon dioxide using hydrogen sulphide (H2S) as a hydrogen source, whereas in higher plants the source of hydrogen is water. 6CO2 +12H2S ![]() C6H12O6 + 6H2O + 12S ... (1)
C6H12O6 + 6H2O + 12S ... (1)
6CO2 +12H2O![]() C6H12O6 + 6H2O + 6O2 ... (2)
C6H12O6 + 6H2O + 6O2 ... (2)
In both the above processes carbon dioxide is used but no oxygen is evolved by sulphur bacteria, while on the other hand, it is evolved in higher plants.
It confirms that source of oxygen is water and not carbon dioxide as was believed earlier.
On this basis Van Niel gave following generalized equation for photosynthesis in higher plants and bacteria:
CO2 + 2H2A ![]() (CH2O) + H2O + 2A
(CH2O) + H2O + 2A
In this presentation, H2A represents a hydrogen donor; in green plants this is water (H2O) and in sulphur bacteria it is hydrogen sulphide (H2S).
The action of light might be to produce a reducing entity [H] and an oxidising entity [O] from water.
The views of Van Niel were further confirmed by Ruben, Hassid and Kamen (1941) using radioactive isotope of oxygen (O18).
When they used water (containing radioactive oxygen, H2O18), (O18) was evolved.
It thus confirms Van Niel's observation that the source of oxygen evolved in photosynthesis is water.
Equation (2) provides the most adequate explanation of photosynthetic process, since it shows that oxygen evolved during photosynthesis comes from water.
(4) Red Drop:
Robert Emerson measured the efficiency of photosynthesis of Chlorella cells in monochromatic light (i.e. only one wavelength of light at a time).
They observed that the quantum yield of photosynthesis decreased very sharply towards 680nm and longer wavelengths.
Since this decrease in quantum yield is observed at the far-red region of the spectrum, hence it is called red drop.
Quantum requirement: It is the number of light quanta needed for production of one O2 molecule or reduction of one CO2 molecule. It is 8.
Quantum yield: It is the number of oxygen molecules evolved per light quanta absorbed. Two light quanta are required to move one electron. Such four electrons are involved in the evolution of one molecule of oxygen. Eight quanta are, therefore required for evolution of one oxygen molecule. Thus quantum yields or number of oxygen molecules evolved per light quanta is 1/8.
Enhancement Effect
Emerson found that when light of shorter wavelengths was used superimposed with the light of long wavelengths, the rate of photosynthesis was greater (25% more) than could be expected from adding the rates found when either light was given alone.
Thus both light beams had a synergistic effect i.e. one helped the other.
It leads to the conclusion that two group of pigments, one absorbing in lower range and other absorbing in higher range of wavelengths are involved in photosynthesis and are called pigment systems.
(5) Two pigment systems :
It is a group of pigment molecules having chlorophyll a, chlorophyll b, carotenoids etc.
One molecule of chlorophyll a functions as a reaction centre or trapping centre and others act as light harvesting complex (LHC) or antenna molecules.
These antenna molecules collect light energy and transfer this energy (by inductive resonance process) to reaction centre where primary photochemical act occurs (quantum conversion) i.e. absorption of a photon and release of an electron.
Mechanism of Photosynthesis
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Mechanism of Photosynthesis
LIGHT REACTION OR HILL REACTION
It occurs in lamellar part of chloroplast an antasomes are units of light reaction. It includes following main processes.
(1) Light and its absorption :
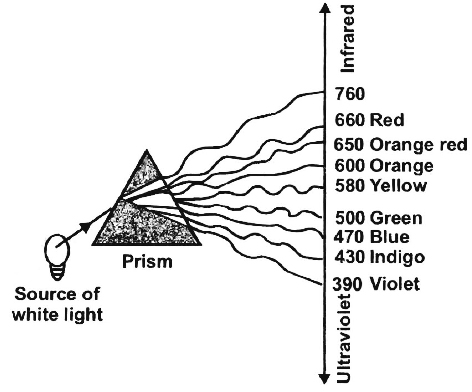
White light, as it reaches us from the sun, is composed of different wavelengths.
When passed through a glass prism it resolves into seven colours-violet, indigo, blue, green, yellow, orange and red.
This band of seven colours constitutes the visible spectrum.
Visible light consists of radiations having a wavelength between 390-760 nm.
Part of spectrum used in photosynthesis has a wavelength between 400 -700 nm and is called PAR i.e., photosynthetically active radiation.
In the range of visible spectrum violet bas the shortest wavelength (390-430 nm) and red the longest (660-760nm).
Wavelength longer than 760 nm (infra red radiation) and shorter than 390 nm (ultraviolet radiation) also exist but these are invisible.
Only a small portion of visible spectrum reaction the earth is utilized in photosynthesis.
Chlorophyll - the green pigment of leaves, absorbs mainly the blue 470 -500 nm) and red (660 -760 nm) wavelengths.
It reflects the green wavelengths (500-580 nm) and that is why the leaves appear green in colour.
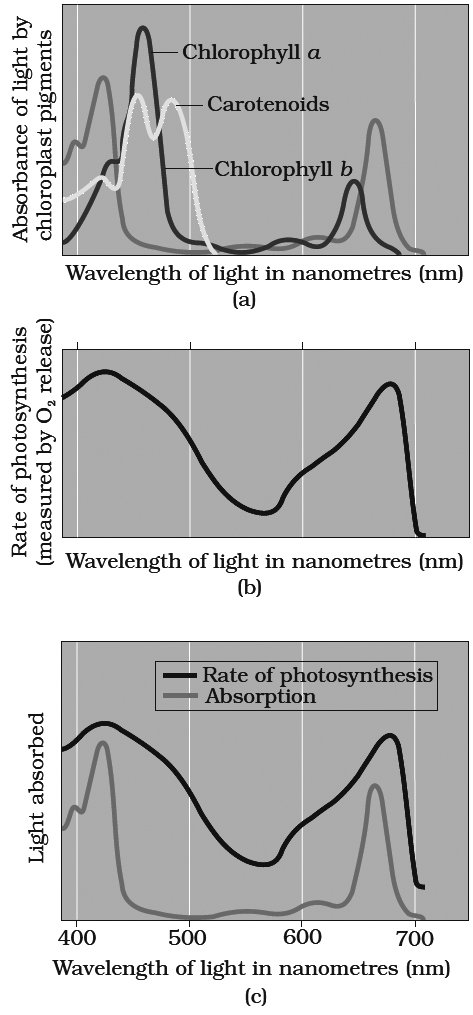
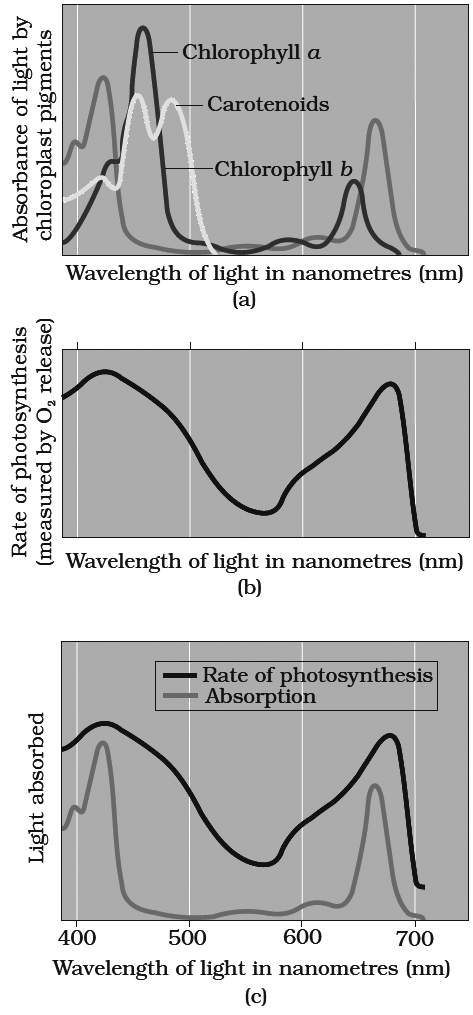
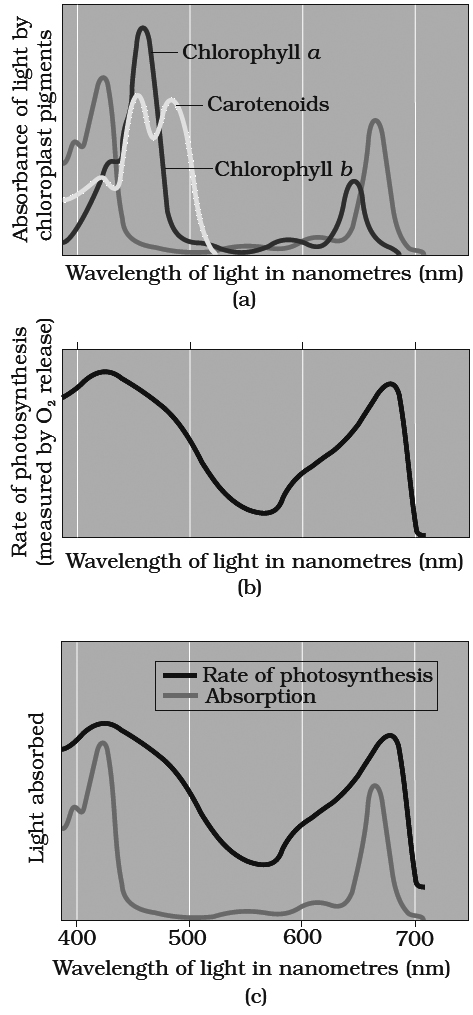
Graph (a) showing absorption spectra of chlorophylls a, b and carotenoids.
Graph (b) showing action spectrum of photosynthesis. Graph (c) showing action spectrum of
photosynthesis superimposed on absorption spectrum of chlorophyll a
Absorption and Action Spectra
Different plant pigments absorb only certain wave lengths of light and these wavelengths are not absorbed at the same rate.
A curve obtained by plotting the amount of absorption of different wavelengths of light by a particular pigment is called absorption spectrum.
An action spectrum is a curve showing the effectiveness of different wavelengths of light in stimulating the process being investigated.
The effectiveness of different wavelengths of light on photosynthesis is measured and plotted by quantum yield or amount of action (expressed through CO2 reduction, O2 evolution etc.) Action spectrum of photosynthetic pigments was studied by Engleman on Cladophora and Spirogyra for the first time.
(2) Excitation of chlorophyll by light :
Each molecule of chlorophyll is composed of many atoms of hydrogen, oxygen, carbon, nitrogen, etc. Negatively charged electrons of different energies revolve in specific orbits around positively charged nucleus of the atom.
Each electron possesses a definite amount of energy depending upon its distance from the nucleus.
This state of electron is known as ground state -a condition of maximum stability.
If a sufficiently energetic photon of light strikes one of the electrons, the electron acquires the energy of photon (quantum) and jumps to another orbit of higher energy.
This is said to be the excited state of the electron.
It is an unstable state and the electron stays in this state for only a very short time (10–5 – 10–9 second).
As the excited electron returns back to its ground state, the trapped energy may be emitted as heat, light or as fluorescence.
In green plants, however, the energy emitted by the excited chlorophyll molecule is trapped and stored in the form of chemical energy (ATP) rather than being lost simply as light or heat.
If the excited electron returns to its ground state in one step, the energy stored in it would be released in one large dose and may not be fully converted into chemical energy.
Therefore, the excited electron, as it returns to its ground state, is passed on to a number of acceptors so that the maximum amount of released energy is converted into chemical energy.
Concept Builder
Photoluminescence :
It is the property of reradiation of absorbed energy. The reradiated energy is usually or longer wavelength. It of two types.
(i) Fluorescence : It is immediate reradiation of fight energy. It stops immediately after the withdrawal of source of illumination. All types of photosynthetic pigments are fluorescent. Most of the fluorescence produced by green parts are due to chlorophyll-a.
Isolated chlorophyll 'a' when dissolved in organic solvents and exposed to light emits red colour immediately. It is called fluorescence; and this quality is called kutusky effect. It is, infact, a manifestation of loss of light energy absorbed in excess. Actually, when the absorbed energy is not utilised for a purpose (phosphorylation), it is emitted out as radiations of longer wavelength. In green plants, of course, this is utilised to synthesise chemical energy (ATP).
(ii) Phosphorescence: The delayed re-radiation of absorbed energy is called phosphorescence.
Non - Cyclic Photophosphorylation
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Non-cyclic photophosphorylation:
It involves photosystem I as well as photosystem II. Both these systems absorb light photons. For convenience, the non-cyclic photophosphorylation is described here in following steps:
(i) When P680 absorbs a photon of red light, it gives out an electron, and thus becomes positively charged. This electron from P680 passes to P700 through a series of electron carriers, like Phaeophytin (Primary acceptor), plastoquinone, cytochrome b6-f complex, plastocyanin.
This movement of electron is downhill in terms of redox potential scale.
In this series, enough energy is released when the electron is transferred from PQ to cytochrome b6-f complex. It is utilized to synthesize ATP.
(ii) Simultaneously, electron in the reaction centre of PS I also get excited when they receive red light of 700 nm and these electrons are transferred to another receptor having high redox potential. These electrons then move downhill again, this time to a molecule of NADP+. This addition of electron and a proton, reduce it to NADPH + H+.
1.The process requires an external electron donor.
2.It is connected with photolysis of water and liberation of oxygen.
3.It is not only connected with ATP synthesis, but also with production of NADPH.
4.Non-cydic photophosphorylation takes place under optimum light, aerobic conditions and in the presence of carbon dioxide.
5.The system is connected with CO2 fixation.
6.It occurs in the granal thylakoids.
7.DCMU inhibits noncyclic photophospho- rylation.
Splitting of water
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Splitting of water
(iii) Photolysis of water and replacement of ejected electron of P680. Breakdown of water molecules under the influence of light is known as photolysis.
The positively charged P680 molecule (after having given an electron) exerts a strong pull on the water molecule, splitting it into H+ and OH– ions. This is called photolysis of water. It requires presence of OEC (oxygen evolving complex) enzyme, Cl– and Mn++ ions as catalysts (Ca+2 ions control their concentrations).
4H2O![]() 4(OH)– + 4H+ ...(i)
4(OH)– + 4H+ ...(i)
4(OH)– ![]() 4e– + 4OH ...(ii)
4e– + 4OH ...(ii)
The OH– donate their electrons to oxidised P680 so that it once again becomes functional.
The (OH)– radical obtained in equation (ii) above eventually forms water and liberates oxygen.
4(OH)![]() 2H2O + O2 ...(iii)
2H2O + O2 ...(iii)
Hydrogen ions (4H+) formed by the photolysis of water {equation (i)}, accumulate within the lumen of thylakoids.
Redox potential is the tendency of an atom or molecule having low redox potential to lose electrons (electron donors), while those with high redox potential to accept electron (gain electrons).
Hence, electrons move from substances having low redox potentials to those having high redox potentials.
Assimilatory power: NADPH + H+ and ATP produced in light reaction constitute assimilatory power, NADPH + H+ produced in light reaction is called as reducing power.
Chemiosmotic Hypothesis
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Chemiosmotic Hypothesis
Generation of ATP in light reaction is called photophosphorylation.
Chemiosmotic hypothesis was first explained by P. Mitchell.
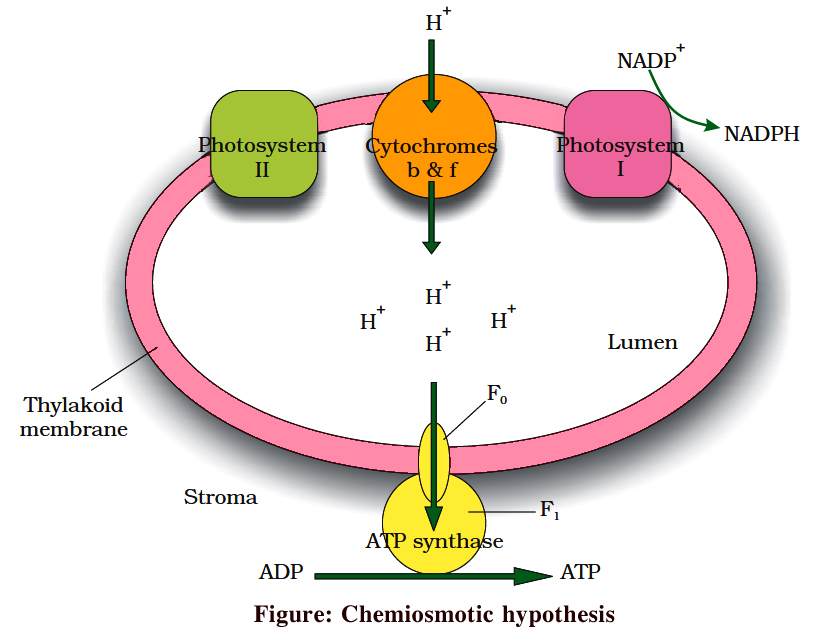
As a result of (a) photolysis towards thylakoid lumen, (b) quinone pump, which takes H+ from stroma to lumen, and (c) NADP+ reductase reaction towards stroma; H+ concentration in thylakoid lumen increases by 1000-2000 times and as a result a proton motive force develops.
This force is responsible for ATP synthesis on the head of CF0 – CF1 particle plugged on thylakoid membrane. The CF0 part forms facilitated channel for proton diffusion towards stroma. Movement of 3H+ is involved in the synthesis of 1 ATP.
Where are ATP and NADPH used ?
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
ATP and NADPH used
DARK REACTION / PCR OR BLACKMAN REACTION
The light independent or dark reaction of photosynthesis reduces the carbon dioxide to glucose.
These reactions take place in the stroma of chloroplast where light independent enzymes are present.
Although this process does not require light, it depends upon the products of light reaction of photosynthesis.
The energy required for the reduction of carbon dioxide is derived from ATP and hydrogen from NADPH (formed in the light reaction of photosynthesis).
The dark reaction occurs through one of the following three methods under different conditions by various plants:
(1) Calvin cycle or C3 cycle
(2) Hatch and Slack cycle
(3) CAM cycle
(1)CALVIN CYCLE or C3 CYCLE
This process comprises a series of reactions controlled by enzymes.
The sequence of these reactions were determined in Chlorella and Scenedesmus by Calvin, Benson and Bassham using radioactive carbon 14C, and techniques like chromatography and autoradiography.
Therefore, it is also known as Calvin cycle or Calvin-Benson cycle.
First product of CO2 fixation is a 3C compound called 3-phosphoglyceric acid (PGA).
This cycle occurs in all photosynthetic plants; whether they have C3 or C4 or CAM pathways. The enzyme for CO2 fixation is RuBisCO.
It is a large enzyme found in stroma.
It is most abundant protein on earth and constitutes 16% of chloroplastic protein.
It shows bifunctional nature having both carboxylase and oxygenase activity.
The dark reaction is also known as Blackman's reaction.
The whole reaction can be studied in three parts:
(i) Carboxylation (Acceptance of CO2 by RuBP i.e., CO2 acceptor -it is a 5C compound),
(ii) Reduction, (iii) Regeneration of RuBP.
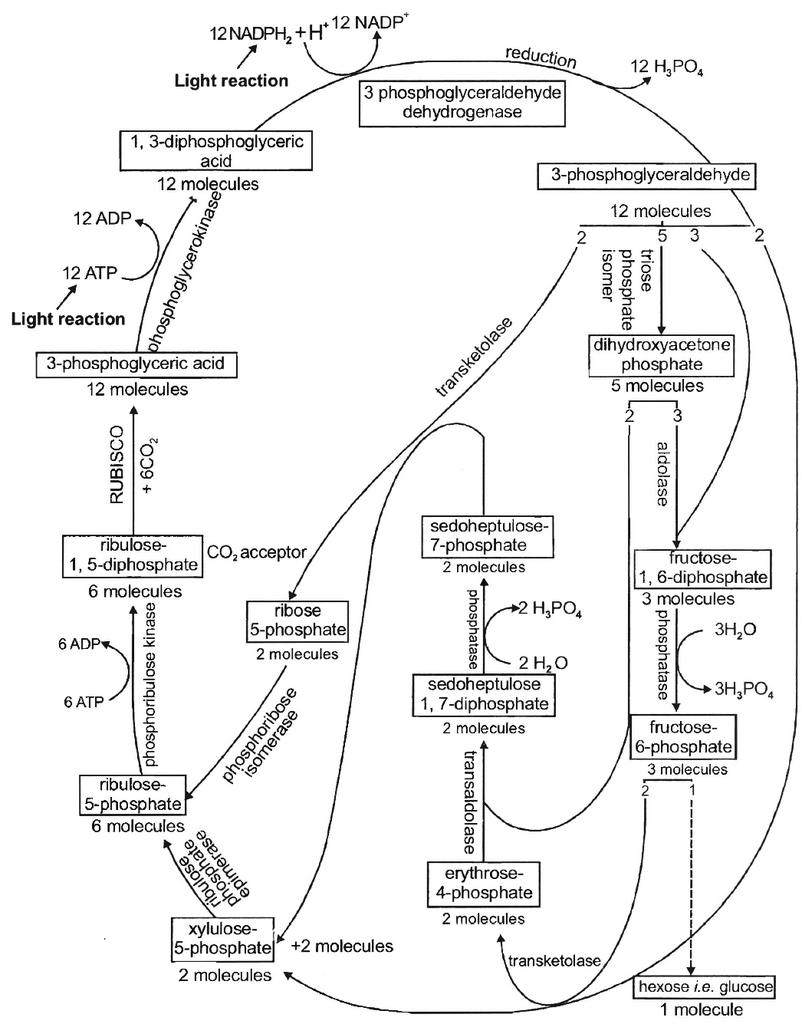
18 ATP and 12 NADPH + H+ are used for the formation of a molecule of glucose (or to reduce 6CO2).
During this process, 264 grams of CO2 and 216 grams of water are used for producing 108 grams of water and 192 grams of O2.
End product of photosynthesis. Hexose sugars are considered as the end products of photosynthesis. Generally, hexose sugar is stored in the form of starch, a long and branched chain of repeating units of simple sugars (usually glucose). Besides, under certain circumstances phosphoglyceraldehyde may also directly or indirectly give rise to fats and proteins in addition to hexose sugars.
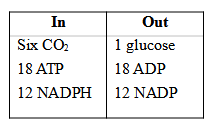
(1) carboxylation, during which CO2 combines with ribulose-1, 5-bisphosphate;
(2) reduction, during which carbohydrate is formed at the expense of the
photochemically made ATP and NADPH; and
(3) regeneration during which the CO2 acceptor ribulose 1, 5-bisphosphate is formed again so that the cycle continues.
The C4 Pathway {Hatch and Slack Pathway}
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
The C4 Pathway {Hatch and Slack Pathway}
OR CO-OPERATIVE PHOTOSYNTHESIS
Most of the plants are known as C3 plants because the first product of CO2 fixation is a 3 C compound, phosphoglyceric acid.
But C3 cycle (or Calvin cycle) is not the only way in plants to fix CO2, H.P. Kortschak and C.E. Hartt (1965) found that in sugarcane (a tropical plant), leaves removed CO2 more efficiently from the atmosphere; and the first products of photosynthesis were acids containing 4-carbon atoms (e.g., malic, oxaloacetic and aspartic acid), rather than the 3C-acid PGA.
Since then, the same has been found true for many important tropical plants including monocots (like maize, Sorghum and Eleusine) as well as dicots (like Amaranthus and Euphorbia sp.).
These plants are called C4 plants. In 1966, two Australian scientists, Hatch and Slack showed that C4 plants are much more efficient in CO2 utilization than C3 plants.
Such plants do not show photorespiration.
In 1967, Hatch and Slack explained the manner of CO2 fixation and reduction in such plants.
The new carbon pathway in C4 plants is called Hatch-Slack Pathway.
The leaves of C4 plants show Kranz anatomy, means there vascular bundles are surrounded by two rings of cells:
(i) Bundle sheath cells : Thick walled, packed tightly all around the vascular bundle, plastids centripetally arranged, lack grana, contain starch grains, absence of PS II, CO2 released by malic acid is fixed by RuBP. Calvin cycle enzymes are present in stroma.
(ii) Mesophyll cells : Concentrically arranged, form most of the leaf tissues; plastids well developed, grana large and well developed; PS II present, CO2 accepted by PEP, CO2 fixation occurs in cytoplasm; Calvin cycle enzymes are absent.
In C4 plants, CO2 is first accepted in the mesophyll cells by phosphoenol pyruvic acid (a highly energetic Cintermediate of glycolysis) using the enzyme PEPCO/PEPCase.
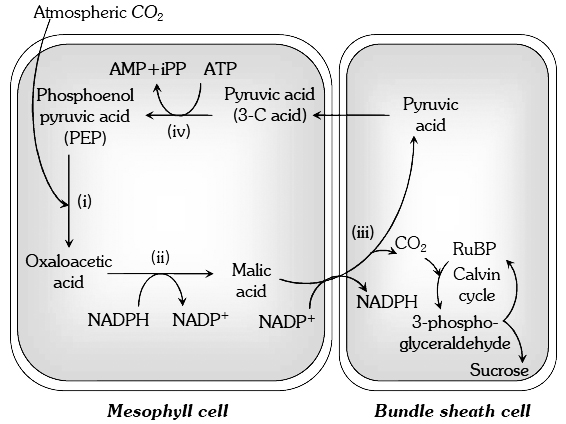
and bundle sheath cells of C4 plants
This forms a C4 compound oxaloacetic acid.
It transforms into another 4 C compound malic acid.
The malic acid now diffuses to the bundle sheath cells where it gives off CO2 It is accepted by ribulose diphosphate and refixed into hexose sugars through Calvin cycle.
The malic acid, after giving off CO2 convert into pyruvic acid and returns to mesophyll cells. It is then converted to phosphoenolpyruvate by using ATP.
This involves formation of AMP (adenosine monophosphate) instead of ADP.
Hence, regeneration of ATP from AMP requires 2 ATP each (12 ATP for formation of 6 molecules of phosphoenolpyruvate, as the 6CO2 molecules are to be fixed).
Therefore, C4 pathway requires 12 additional ATP or total 30 ATP (18 ATP in C3 cycle + 12 additional ATP).
An interesting aspect of C4 cycle is that, it shows a division of labour between two kinds of parenchyma cells showing chloroplast dimorphism -mesophyll chloroplasts act as traps of CO2 which is then (in the form of C4 acids) shuttled to bundle sheath plastids where finally the CO2 is reduced to sugars.
C4 plants are more efficient in picking up CO2 and tolerating drought conditions.
CO2 compensation point C4 is low (0-10 ppm) in C4 plants (as compared to 25-100 ppm) for C3 plants. Photorespiration is absent in C4 plants, because they have a mechanism to concentrate CO2 at enzyme site.
Some major differences between C3 and C4 plants

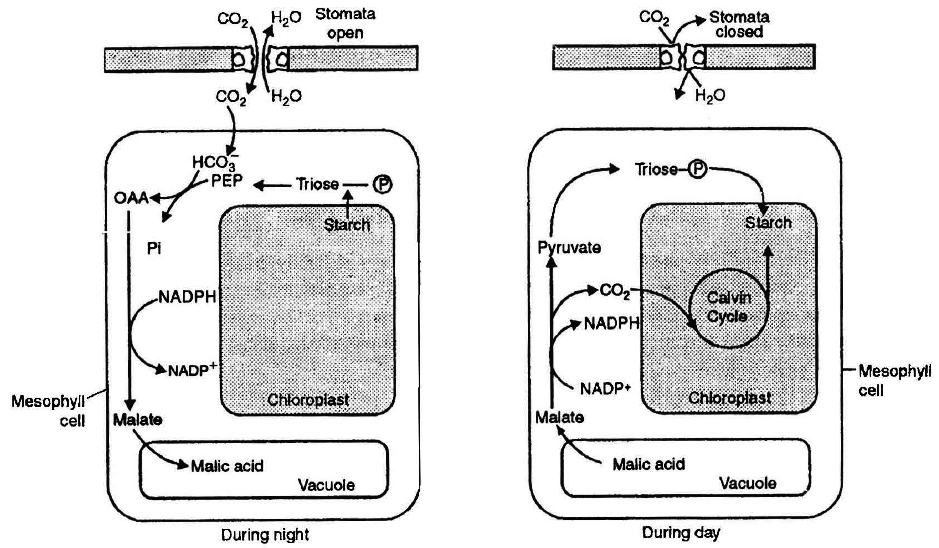
(3) CRASSULACEAN ACID METABOLISM (CAM) (Diurnal acid Cycle
(i) The leaves of plants belonging to Crassulaceae family like Kalanchoe and Sedum undergo crassulacean acid metabolism and such plants are called CAM plants.
(ii) All CAM plants are succulent in habit, including other plants like Orchids, pineapple, certain cactus etc. In these plants, stomata remains open in night and closed during the day (Scotoactive stomata).
(iii) CO2 is fixed during night (dark) to OAA using PEP carboxylase. This CO2 comes from respiration (breakdown of starch) and also from the atmosphere. Malic acid gets stored in vacuoles.
(iv) The CAM plants also contain the enzymes of Calvin cycle. During day time, malic acid breaks into pyruvate and CO2, While CO2 enters the calvin cycle, pyruvate is used up to regenerate PEP.
(v) The succulents, therefore synthesize plenty of organic acids from CO2 during night (when stomata are open) and plenty of carbohydrates during the day (when stomata are closed)(vi) Like Calvin cycle, CAM cycle also operates in the mesophyll ceil. None of these have shown chloroplast dimorphism as is found in C4 plants.
(vii) t should be remembered that the slow growing desert succulents exhibiting CAM cycle have the slowest photosynthetic rate, while the species possessing C4 pathway possess the highest rates.
(viii) Thus CAM plants are although not as efficient as C4 plants, they are definitely better suited to the adverse conditions (i.e., conditions of extreme desiccation).
Photorespiration
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
PHOTORESPIRATION (C2 Cycle or Glycolate Metabolism)
Term given by (Decker and Tio). Photorespiration is a process which involves oxidation of organic compounds in plants by oxygen in the presence of light.
Like ordinary respiration, this process also releases carbon from organic compounds in the form of carbon dioxide but does not produce ATP.
Thus, apparently it seems to be a wasteful process, but it must have some functions which are still unknown.
The process occurs in C3 plants only.
There are two oxygen consuming reactions in this process.
First oxidation occur in chloroplast and second oxidation occur in peroxisome.
Photorespiration occurs usually when there is high concentration of oxygen.
Under such circumstances RuBP carboxylase (or Rubisco), the enzyme that joins carbon dioxide to RuBP, functions as oxygenase.
As a result, oxygen, instead of carbon dioxide, gets attached to the binding site of the enzyme (i.e. RuBP).
This reaction is competitive.
It is the relative concentration of O2 and CO2 that determines which of the two will bind to the enzyme.
On oxidation, RuBP releases one molecule of C3 compound.
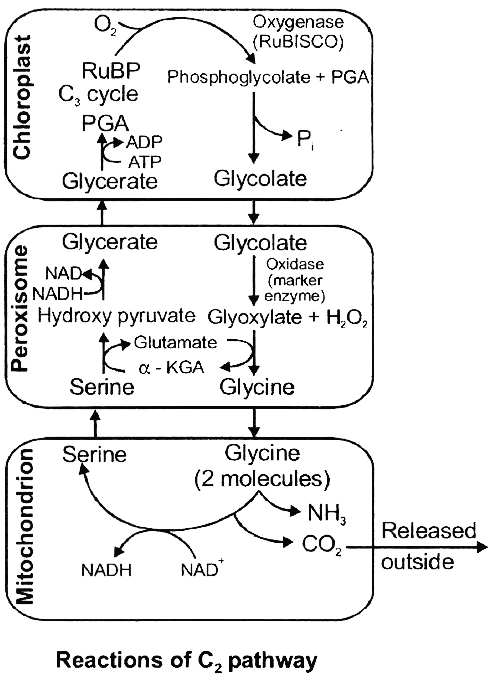
PGA (which enters C3 cycle) and one molecule of a C2 compound phosphoglycolate.
The latter almost immediately changes into glycolate.
The glycolate leaves the chloroplast and enters a membrane bound sac of enzymes called peroxisome.
Here the glycolate is oxidised into glyoxylate which is aminated into glycine.
Further condensation of glycine takes place inside the mitochondria where two molecules of glycine interact and give rise to a molecule of serine and carbon dioxide each.
The CO2 is then released from mitochondria.
Serine regenerates PGA. With increase in temperature, high light intensity and oxygen concentration, affinity of Rubisco for CO2 decreases and its affinity for oxygen increases.
Thus, a rise in temperature means more loss of photosynthetically fixed carbon by photorespiration.
It reduces potential yield of plants growing in the tropics by 30-40%. Upto 1/4th of the photosynthetically fixed CO2 may be lost by this pathway.
Factors affecting Photosynthesis
- Books Name
- ACME SMART COACHING Biology Book
- Publication
- ACME SMART PUBLICATION
- Course
- CBSE Class 11
- Subject
- Biology
Factors affecting Photosynthesis
Photosynthesis occurs under the influence of many factors, both internal and external.
When several factors affect any (bio)chemical process, Blackman's (1905) law of limiting factors comes into effect.
Blackman's Law of Limiting Factors
According to this law "When a process is conditioned as to its rapidity by a number of separate factors, the rate of the process is limited by the pace of the slowest factor."
In other words. "If more than one factors affect a chemical process then its rate will be determined by the factor which is nearest to its minimal value".
It is the factor which directly affects the rate if its quantity is changed.
A limiting factor is that factor which is deficient to such an extent that increase in its value directly increases the rate of the process.
A. External factors affecting photosynthesis
(1) Light :
It is an essential factor for photosynthesis. It affects the rate of photosynthesis in three ways.
I. Light Intensity: It affects the rate of photosynthesis. Optimum value of light intensity is different for different plants.
There are two categories of plants :
(a) Sciophytes : They grow under diffuse light, e.g., Oxalis etc.
(b) Heliophytes: They grow in direct sunlight, e.g., Dalbergia etc.
(i) The optimum light intensity for sciophytes is 10% of full summer sunlight.
(ii) The-optimum light intensity for heliophytes is 50-70% of full summer sunlight.
(iii) Optimum for C4 plants is more than full summer sunlight. e.g., Maize, sugarcane.Optimum light in ensity is also called saturation point.
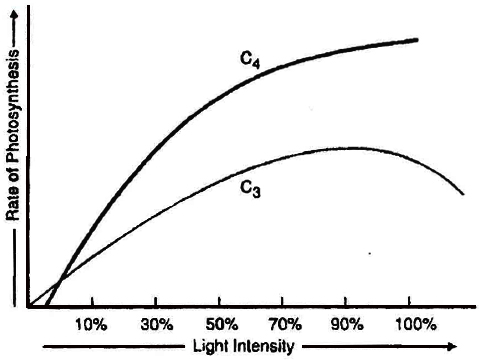
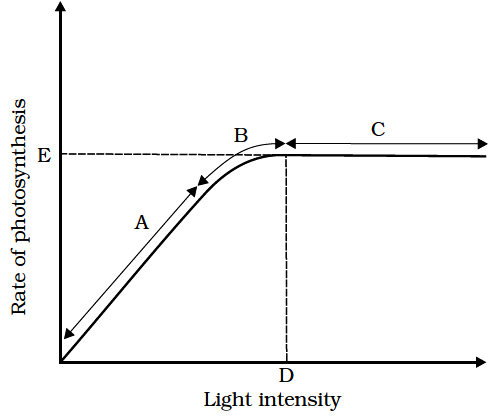
(iv) As the light intensity decreases rate of photosynthesis also decreases. The point in light intensity where there is no apparent exchange of gases between the environment and the photosynthetic organ is called light compensation point. Increase in light intensity from compensation point to saturation point increases the rate of photosynthesis. Light intensity above the saturation point decreases the rate of photosynthesis, this effect is called solarisation. This includes photoinhibition and photo-oxidation.
II. Light quality : Light between 400-700nm wavelength constitute the photosynthetically active radiation or PAR. Maximum photosynthesis takes place when whole white light is available to the plant. Maximum photosynthesis takes place in red and blue light and minimum photosynthesis takes place in green light.
III. Duration of light: Light duration does not affect the rate of photosynthesis, but it affects the total photosynthesis.
(2) Carbon dioxide:
Carbon dioxide concentration is the major factor influencing the rate of photosynthesis.
The CO2 concentration is very low in the atmosphere (between 0.03 and 0.04 percent).
This level of CO2 is far below the requirement for optimum photosynthesis.
Thus, the rate of photosynthesis could be increased several times by increasing CO2 concentration to about 0.05%.
At current level of CO2 concentration the rate of photosynthesis is optimum for C4 plants and lower for C3 plants.
If CO2 concentration increases then only C3 plant would be benefitted. A stage in CO2 concentration when there is no net absorption of CO2 by illuminated plant organ is called CO2 compensation point. At this point, photosynthesis rate is equal to respiration rate. CO2 compensation point for C3 plant is 25-100 ppm and for C4 plant is 0 -10 ppm.
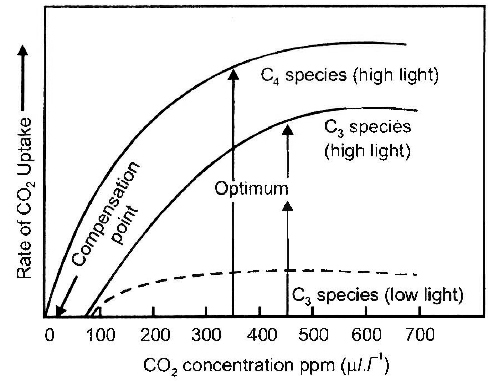
C4 plants show saturation at about 360 ppm, while C3 plants shows this beyond 450 ppm. Thus current CO2 concentration is limiting for C3 plants.
Very high CO2 concentration causes the stomata to close. This inhibits exchange of gases and as a result the photosynthetic rate decreases.
(3) Temperature:
The dark reactions are controlled enzymatically in stroma, hence are temperature regulated.
Optimum value of temperature for photosynthesis in C3 plant is 20-25°C and for C3 plants is 30º–45ºC) The minimum temperature at which most plant starts photosynthesis is 0-5°C. It is however as low as –35°C for some gymnosperms.
(i) Maximum temperature at which photosynthesis can occur is 50-55°C for desert plants (Opuntia) and 70-75°C for hot spring algae.
(ii) Tropical plants have a higher temperature optimum than the plants of temperate climate.
(iii) In C4 plants, pyruvate phosphate dikinase enzyme is sensitive to low temperature, while in C3 plants affinity of RuBisCO for CO2 decreases at high temperature.
(iv) Q10 values for photosynthesis is 2.
(4) Water:
Photosynthetic process utilizes less than 1% of the water absorbed by a plant, hence, it is rarely a limiting factor in photosynthesis.
But water scarcity affects photosynthesis indirectly.
It has been observed in several experiments that the rate of photosynthesis decreases drastically if water supply is withheld for sometime.
Water stress can cause stomatal closing, wilting of leaves and reduced metabolic activities in leave.(5) Oxygen.
Increase in O2 concentration decreases the rate of photosynthesis in C3 plants.
This phenomenon of the reduction of photosynthesis by O2 was first discovered by Warburg using an alga called Chlorella and is called as Warburg's effect.Oxygen is inhibitory because:
(i) It favours photorespiration.
(ii) It destroys the activated state of chlorophyll.
(6) Mineral nutrient.
Few mineral elements are directly involved in photosynthesis. Mg is a component of chlorophyll. Mn, Ca and Cl are essential for photolysis of water.
B. Internal factors affecting photosynthesis
Photosynthesis is under the influence of several internal (plant) factors.
The plant factors include the number, size, age and orientation of leaves, mesophyll cells and chloroplasts, internal CO2 concentration and the amount of chlorophyll.
The plant or internal factors are dependent on the genetic predisposition and the growth of the plant.
(1) Chlorophyll.
Of the internal factors, chlorophyll is the most important because light energy is trapped by only this substance.
There is no photosynthesis in the absence of chlorophyll.
The non green parts of variegated leaves (e.g., Croton), therefore, do not have starch.
Photosynthetic number or assimilation number shows a relationship between the chlorophyll and photosynthesis.
It is the amount of carbon dioxide (in gms) assimilated by one gram of chlorophyll in an hour.
Emerson (1929) observed a direct relationship between the chlorophyll content of a leaf and the rate of photosynthesis.
If all other factors are favourable, increased chlorophyll leads to an increase in photosynthesis.
(2) Photosynthetic products.
With the accumulation of the end products of photosynthesis in mesophyll cells, there is decrease in their photosynthetic rate because concentration of these products in the cells increases the rate of respiration.
Concept Builder
1.'Photosynthesis is an anabolic, endergonic; oxidation-reduction process.
2.Destruction of chlorophyll due to, high light intensity is called solarisation.
3.In C3 plants, more CO2 is released in light than in dark due to photoresprration.
4.Maximum absorption peak of, chlorophyll a and chlorophyll b is in blue light.
5.Chloride ions stimulate quick release of electrons from water.
6.Pheophytin is chlorophyll a ,in which Mg++ is replaced by two hydrogens.
7.Atriplex hastata shows Calvin cycle whereas Atriplex rose us shows Hatch and Slack cycle.
8.The reaction centre in bacteria is B-890 and reducing agent is NADH2, in bacterial photosynthesis, no photolysis of H2O occur, therefore phothesynthesis is an oxygenic and cyclic ETS dominates.
9.Moll's half leaf experiment is used to show that carbon dioxide is essential for photosynthesis.
10.Wilmott's bubbler is used to determine the rate of photosynthesis by counting the O2 bubbles.
